Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to Classics index)
First published in Psychological Review, 41, 134-152.
VI. AUTOMATIC HABIT TRANSFER WHEN THE PRINCIPLE OF FREQUENCY MUST BE OVERRIDDEN
We must now consider the question of how the tendency to take the long path, which is clearly dominant with many of the animals in Valentine's experiment at the beginning of the 'critical runs,' i.e., that portion of the practice in which the door at E was open throughout each trial, finally gives place to a dominance of the tendency to take the short path. We have shown in the preceding section how occasional choices of the shorter path might be expected to take place through the influence of chance sensory disturbances. But in many cases, at least, the external inhibitions at E would be of relatively infrequent occurrence and the proportion of long-path to short-path choices would still greatly favor the choice of the longer path. Suppose, for example, that we assume only one short choice out of six. A naïve interpretation of the principle of frequency would demand that such a ratio not only would not increase the proportion of short-path choices but that the short-path choices should steadily decrease until they would entirely disappear. Such an interpretation of the principle of frequency implicitly assumes that frequency is the only principle operative. It is to be doubted whether anyone has ever utilized this principle under such an assumption. At all events, there is reason in this particular case to believe that at least two other factors are also operating simultaneously and in such a way as to oppose and override it. [14] [p. 135]
The first opposing factor to be adduced in this connection is the empirical principle discovered in conditioned reaction experiments that the restoration of an experimentally extinguished tendency to action is much more rapid than was its original acquisition. Thus Hilgard and Marquis (9) report that the restoration of an experimentally extinguished lid reaction recovered in the case of three dogs in approximately one-fifth, one-fourth, and two-fifths respectively of the numbers of reinforcements originally required to produce the same increase in strength of the tendency. If we may take the middle of these three values as a first rough approximation and apply it tentatively to Valentine's rats, this principle alone would allow the animals to take the long path three times out of four and still have a substantial margin of practice effects in favor of the shorter path.
A second principle which presumably contributes to the same end is that of the goal gradient (16). Let us assume in the interest of simplicity of exposition that increase in strength of excitatory tendency per repetition within a moderate range is constant under given experimental conditions. Now, from the choice point E in Valentine's maze, the long path is nearly four times as great as the short one. Turning to the table of hypothetical goal gradient values (16, p. 32) and giving the short and the long paths distance values of 10 and 38 respectively, it is easy to see that one repetition of the short path will strengthen the tendency to repeat that act on a subsequent occasion much more at point E than one repetition of the long path will strengthen the latter tendency at the same point. Under these assumptions the short path would increase in strength per trial in the proportion of 4.28 whereas the long path would increase in strength per trial in the proportion of only .96. Thus, by this principle alone the short path would ultimately become dominant, even though it were chosen only a fourth as often as the longer one. [15] This pro- [p. 136] portion would vary with the absolute size of the maze, but the general principle ought to hold if the goal gradient hypothesis is sound.
We do not yet know enough about the two processes just considered to be able to say whether they would combine in an additive or a multiplicative manner, but it is evident that they would summate in some way. The nature of the relationship would seem to favor a multiplicative mode of combination. If that were true, the above numerical assumptions would lead to the expectation that an ultimate giving up of the blind alley in Valentine's experiment during the 'critical' runs would still take place even if disinhibition should occur on the average in only one trial out of ten or twelve. By similar reasoning, the more frequently disinhibition takes place at point E, in this stage of the learning process, the more rapidly the blind will be eliminated. On the other hand it is conceivable that distractions might be so far eliminated that occasional animals would never make the correction at all. From this general angle may possibly be explained the stubborn cases of 'fixation' which have puzzled many experimenters; it may be that certain animals, by constitution or the accidents of life, are relatively immune to disinhibition.
VII. THE PREDILECTION OF ANIMALS FOR ENTERING GOALWARD-POINTING BLINDS
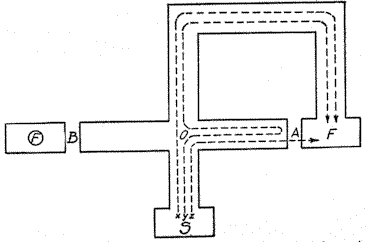
We come now to the consideration of the behavior of the rat in an ordinary single path maze such as that shown in Fig. 13. Experimentalists frequently report two characteristic phases in the process of a hungry rat learning a maze. The first is a slow, hesitant groping until the food is found. Soon after, the behavior of the rat changes markedly: he goes about his searching in a businesslike way, as if he were 'going somewhere' (6). The same thing is often expressed by saying that the rat has become oriented. It will be recalled that according to the hypothesis here being presented, orientation itself consists in the anticipatory goal reaction (rG) becoming conditioned to the external stimulus complex S1' (Figs. 10 and 11). But we have already seen that when [p. 137] this takes place the practice effect of traversing the path by which the food ( F ) was reached is at once transferred to all other members of the particular habit family, especially to those which are shorter and therefore occupy a superior position in the hierarchy. It follows, provided the animal
Fig. 13 Conventional maze to represent the predilection of animals for entering blind alleys which point in the direction of the place where food is obtained. The F in the circle at the left of B represents food inaccessible to the animal but placed in such a position as to give the same olfactory cues at the choice point O as that which is ultimately eaten. The broken lines indicate three conventional paths; path Z represents the path of the preferred member of the habit-family hierarchy which the animal would take if the blind end of alley A did not prevent. It is to be noted that this maze would not be suitable to test experimentally the existence of the tendency to enter goal-pointing blinds because it would also evoke the tendency to make an anticipatory right turn (22, p. 30).
cannot see the end of the alley in question, that every blind is the beginning of a path to the goal. It is obvious that blinds like A , Fig. 13, pointing in the direction of the goal, are the beginnings of paths which, if completed, would be among the shorter ones of the animal's habit family and hence are to be preferred to the longer, more roundabout path, X . It accordingly follows at once that animals will show a marked predilection for blinds pointing in the direction of the goal, as contrasted with alleys pointing in the opposite direction. [16] [p. 138]
As a corollary of the foregoing, it follows directly that for blind alleys pointing in the direction of the food box there will be a period in the learning of the maze, i.e., through the acquisition of orientation, during which these errors will progressively increase. Thus we arrive, by a purely deductive procedure, at the paradox that for certain specific portions of the maze an increase in training will make the score worse rather than better. This deduction may easily be checked by appropriate experiment.
This brings us to the second phase of the maze learning process, the elimination of the blind alleys. It is evident from the preceding discussion that this must be to a considerable extent the elimination of the blinds pointing toward the goal, a form of maladaptive behavior inherently characteristic of the habit-family mechanism. It is to the consideration of the details of this process that we must now apply ourselves.
VIII. The Elimination Of Goalward-Pointing Blinds
There is reason to believe that the maladaptive aspect of the habit hierarchy system just referred to, the predilection for entering blinds pointing in the direction of the food box, is corrected by at least two tendencies. Each of these is fairly distinct, and both presumably operate simultaneously.
The first and more basic mechanism is that of the direct effects of the goal gradient (16) which would make path X (Fig. 13) preferable to path Y because the former is shorter. This principle, however, cannot alone account for the final preference of path X, unless there is assumed to be a moderate number of spontaneous choices of the shorter of the two paths during the early practice trials. [17] But the habit-family [p. 139] principle, as applied to paths X and Y, would, by virtue of the principle of transfer, shift the dominance of reaction from Y to Z , once orientation had taken place on the basis of runs in Y , even though not a single run had ever been made by path Z . Thus, the situation is complicated by the necessary implication that, as already noted in the preceding section, the habit-family principle also produces the strong maladaptive tendency to attempt to take path Z, quite apart from any specific practice whatever on this particular path. We must now consider what phenomena should be expected under these circumstances to result from repeated trials.
In order to understand the outcome of this complex situation, it will be necessary to put forward a supplementary hypothesis already referred to and employed in another connection (p. 41, Part I). This may be called the frustration hypothesis. The term frustration is here used to indicate any situation in which an acquired excitatory tendency, and particularly an excitatory tendency located posterior to a chain of such excitatory tendencies making up an appreciable segment of a behavior cycle, is for any reason prevented from evoking its accustomed reaction. The hypothesis is that under such circumstances internal inhibitions will be developed which will manifest substantially the same characteristics as result from the experimental extinction of conditioned reactions . Among the phenomena to be expected on this hypothesis are the gradual cessation of attempts to perform the act which has been frustrated, spontaneous recovery of the tendency with the passage of time, and disinhibition (25, p. 66) .
It will be necessary also to recall in this connection the famous study of Ebbinghaus (4; 7). In that investigation he obtained experimental evidence indicating that in series of nonsense syllables learned by rote, there were set up by the learning process excitatory tendencies not only connecting each syllable with the next but also remote excitatory tendencies between syllables as far apart as seven intervals in the series (4, p. 106). It is the existence of these remote excitatory tendencies that is of special significance in the present connection. [p. 140]
With these principles especially in mind, let us proceed to the consideration of what might logically be expected to take place in the behavior of the rat upon repeated entries into blind alley A of Fig. 13. Suppose that the stimulus and behavior flux of potential path Z , from choice point O to the goal F , be represented by the arbitrary action phases shown in Fig. 14, with the barrier (the end of the blind) falling just
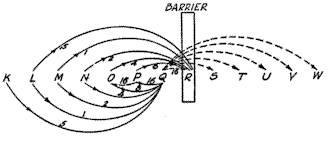
Fig. 14. Diagrammatic representation of some of the immediate and remote excitatory tendencies involved in a typical behavior sequence where an animal in amaze enters a blind alley pointing in the direction of the food box. The letters K, L, M, N, etc., represent both stimuli and reactions at closely spaced intervals in the behavior flux. For purposes of simplifying the exposition, the goal gradient as such is ignored. The gradient of the remote excitatory tendencies of Ebbinghaus is represented conventionally by giving each value one-half the strength of that preceding it. Each point is understood as having six excitatory tendencies emanating from it when considered as a stimulus, and six excitatory tendencies converging upon it when considered as a reaction. Only a few of these could be represented without unduly complicating the figure.
before reaction R. Now, according to the Ebbinghaus hypothesis, there should be an immediate excitatory tendency with a strength of 16 units, say, from Q to R ; a remote excitatory tendency of less strength, say one-half as great (8 units), from P to R ; a second remote excitatory tendency, one-fourth as great (4 units), from O to R, and so on. The excitatory tendencies of this series will yield a total aggregate excitatory tendency of approximately 32 units. It is important to note that because of the habit-family principle there will be at all times, in addition to the tendency of path Z to lead into alley A , a relatively weaker excitatory tendency to go out of alley A by path Y ; Let us assume that this latter and opposing tendency has a strength of 14 units. [p. 141]
But the barrier, by preventing act R from taking place, according to the frustration hypothesis, will produce an inhibition at all points from L to Q , presumably of a strength at each point in proportion to the strength of the original excitatory tendency from each point to R (25, p. 100). Now it is known that when one excitatory tendency suffers extinction, the resulting internal inhibition has a tendency to irradiate, as secondary inhibition, to closely related excitatory tendencies (25, p. 61, p. 171). Thus it would be reasonable to suppose that the frustration of the 8-unit remote excitatory tendency from P to R , together with probable additional inhibitions resulting from the frustration of the excitatory tendency from P to S, P to T, P to U , etc., will spread in such a way as to inhibit to a considerable extent the immediate excitatory tendency from P to Q , and to a progressively lesser extent to the excitatory tendency from O to P , from N to O , from M to N , and so on. Thus there would result from the frustration at R an inhibitory gradient extending backward from R as a focus at which point it presumably will be nearly equal to the inhibitory tendency upon which it is based. This means that there must be a parallel but reversed excitatory gradient extending backward from R , at which point the effective limiting excitatory value would be zero.
Now, by hypothesis, the moment that inhibition has weakened the excitatory tendency to go forward at any point below the assumed level of 14 units, the animal will turn and take path Y out of the blind to the food box F. It is clear that this turning must take place first at point R, since that is the initial focus of frustration at which the inhibition is maximal. Upon repetitions of frustration at successive trials, the inhibition (and so the secondary inhibition) will grow more profound, the gradient at the same time extending farther backward beyond M, L, etc. As a result, the critical excitatory level of 14 units on path X will move backward toward L progressively and as this takes place the point at which turning occurs will also move backward. But when the turning point begins to move backward, e.g., when the turning takes place at Q, this itself will create a [p. 142] new set of immediate and remote frustrations substantially like those which resulted when the barrier prevented act R from taking place, This new set of inhibitions will cause the animal to turn and follow path Y at a still earlier point; the earlier turning will produce another new crop of frustrations, which will result in a still earlier turning, and so on. Thus the animal may be expected, in general, to penetrate the blind alley shorter and shorter distances on successive trials until finally he will not enter it at all, but will proceed directly to the food by way of path X (3; p. 17). [18]
It should be noted that this explanation of the elimination of a blind alley is distinctly related to the important hypothesis put forward by Holmes around 1910 (11, p. 164 ff.). Holmes' principles, however, are inadequate to explain the elimination of a blind alley from a maze because they do not show how the end of an alley can lead to an 'incongruous' act (10). The checking of locomotion or the necessity of turning about at the end of a blind alley only becomes 'incongruous' on the assumption that there is a definite excitatory tendency to go directly through the space occupied by the end of the blind. This is not provided by Holmes' hypothesis. There is no reason to believe, for example, that a rat would develop any inhibition from going into a blind alley during random exploration, for the reason that in such a case there would exist to be frustrated no excitatory tendencies to go anywhere in particular. The principle of the habit-family hierarchy furnishes a clear basis for a definite frustration, and the results from conditioned reflex experiments furnish a principle by which this frustration could be converted into an inhibition without which repetition could hardly produce the elimination. It also affords a deduction of the mode of elimination, i.e., the progressively shortened entrances, a matter of considerable corroborative significance.
IX. EIGHT COROLLARIES INVOLVING BLIND ALLEY ELIMINATING
A number of corollaries flow from the deduction presented in the preceding section:
The first concerns the relative ease of eliminating long vs. short blinds. It has been shown elsewhere (16) that the mechanism of the goal gradient alone would lead to the more ready elimination of long than of short blinds. Since, other things equal, alleys not pointing toward the food box are believed to be eliminated largely by the goal gradient mechanism, it follows that with such alleys hungry animals will eliminate long alleys more readily than short ones.
In the case of alleys pointing in the direction of the goal, however, the principles put forward above lead to a different expectation. Since the elimination is a progressive or step-wise process, it would seem that the deeper the alley, the more steps of a given length it would require to complete the elimination. Consequently long alleys pointing in the direction of the food box should be eliminated less readily than short ones. The third corollary concerns the behavior of the animal between S and 0 , Fig. 13, when he is about to discontinue entering A . Because of the progressive backward movement of the inhibitory gradient, the forward excitatory tendency of path Z should show a perceptible degree of weakening, presumably in the form of reduced speed, for some distance preceding the arrival at the point at which the turning takes place. It would seem logical to expect that as the critical turning point reaches the entrance to the blind (O) this antecedent segment of inhibition should have spread down-ward on path Z towards S. Since inhibitions are known to persist to a certain extent, this should manifest itself by a reduction in the speed of locomotion as the animal approaches O , even on occasions when he has ceased to show any other tendency whatever to enter alley A .
A fourth corollary is dependent on the additional assumption that the dominant principle of the elimination of alleys pointing away from the goal depends upon a rather different [p. 144] principle, i.e., upon the competition at point O (16, p. 34) between the excitatory tendencies to take paths of the X and Y type. If the Y path should triumph on any given occasion, it is difficult to see what, save some scant and rapidly fading inhibitions resulting from the conflict, could prevent the animal from running the whole length of the alley at each entrance, or at least to a point from which the blind end would be visible (17). But since alleys pointing away from the food box are the beginnings of possible, even though non-preferred, paths to the goal, there probably would be some tendency to inhibitory phenomena here also, though it should be weaker than in alleys pointing toward the food box. Accordingly it is believed that if an average were made of the depth of penetration into the two types of alleys on those occasions where a clear tendency to enter appeared, the alleys pointing away from the goal would display the greater mean depth of penetration.
A fifth corollary is that, since the failure to enter alley A is essentially an inhibitory phenomenon, some disturbance like the weak sounding of a buzzer taking place when the animal is on his way between S and O should produce a disinhibition; in which case the animal might be expected again to enter alley A. If a much stronger disturbance should take place it ought also to produce external inhibition of the movements constituting path X, in which case the animal would be likely to cease locomotion altogether. Moreover, upon resumption of locomotion the animal might be expected to go sometimes into alley B , though there would still probably be somewhat of a predilection for entering alley A .
But since the results of both of these types of disturbance are distinctly transitory (25), it may be anticipated with some confidence that after a few minutes of quiet following entrances into the blinds the animal's behavior should show little or no tendency to deviations from its former error less performance.
The fact of spontaneous recovery from experimental extinction demonstrates that inhibitory tendencies disintegrate more rapidly with the elapse of time than do the [p. 145] excitatory tendencies upon which they are based. It seems likely that the principle of the more rapid disintegration of inhibitory tendencies may be rather general, regardless of their origin (25, p. 66). If we are warranted in making this assumption, it follows that the frustrational inhibitions by which goalward-pointing alleys are eliminated would accumulate more slowly if a few hours should intervene between repetitions than if they are immediately consecutive, i.e., other things equal, goalward-pointing alleys will be eliminated less easily by distributed than by massed practice. This principle would be much less active in alleys pointing away from the goal because their elimination is less dependent on inhibition. It follows that with distributed practice there should be relatively a greater difference between the ease of eliminating the two types of alleys than with massed practice, particularly where the alleys appear near the posterior end of the maze. [19]
It is stated (5, p. 365) that caffeine tends to facilitate the acquisition of excitatory tendencies, But to retard the development of inhibitions. In this connection it will be remembered that alleys pointing away from the goal are believed to be eliminated largely on the basis of competing excitatory tendencies, whereas the major factor in the elimination of blinds pointing toward the goal is believed to be inhibitory in nature. If we make the rather dubious assumption that the action of caffeine on the nervous system is as simple as just stated, the above considerations lead to a further corollary: rats learning a maze under the influence of caffeine will eliminate alleys pointing away from the goal more readily [p. 146] than when in the normal condition, But will eliminate alleys pointing toward the goal with greater difficulty than when in the normal condition. Related implications may also be drawn from the alleged action of bromides (5, p. 365).
X. THE CONCEPT OF HABIT-FAMILY HIERARCHIES BASED ON SUBORDINATE GOALS
The above deductions from the concept of the habit-family hierarchy are drawn, for the most part, from the relatively simple situation in which the goal reaction (rG) or some closely related reaction is the dominant factor. There is evidence, however (30; 22; 23), that distinctly subordinate goal reactions also tend to come forward in behavior sequences in a manner substantially similar to goal reactions them-selves, though doubtless less strongly. A large number of experimental phenomena may be deduced, and thus explained, on this assumption by methods analogous to those employed above. Space here is sufficient only to list two or three major type situations.
The first is dependent upon positive transfer on the basis of an rG from a subordinate goal reaction. Here belong Maier's ingenious experiments which indicate that even in the rat the more remote end of a segment of a pathway leading to the goal is able to attract to itself, though weakly, a second segment appropriate in combination to make it possible for the animal to reach the food box (24). The second type is represented concretely by a nice experiment reported by Tolman and Honzik (35; 34). In this study, evidence was obtained which indicates that when an animal encounters a barrier in the segment of a path common to the terminal portion of two alternative paths leading to food, there results an inhibition not only of a previously existent tendency to enter the path traversed on the occasion of the frustration, But of the tendency to enter the other path as well. This phenomenon is explained on the basis of negative transfer, i.e., the transfer to one path of an inhibition acquired in association with the other.
But once the principle of transfer on the basis of sub- [p. 147] ordinate habit families based upon the anticipatory nature of subgoals is recognized as a general proposition, each sub-ordinate goal with its habit family may be conceived as possessing numerous goals of a still lower order, each with a lower-order habit-family hierarchy, and so on. In this way the principle of transfer obviously may be extended down-ward almost indefinitely to even the most minute details of behavior adjustment. On this hypothesis the rich variability, the almost fluid flexibility of behavior characteristic of mammals even in well-practiced action sequences, is a phenomenon not only wholly to be expected from the present point of view, but definitely demanded by it.
XI. SUMMARY
We may now summarize our conclusions in brief dogmatic form. The convergent excitatory mechanism is capable of mediating the transfer of reactions from one stimulus situation to another where the two stimulus situations externally may contain no common element whatever. When the divergent and the convergent mechanisms are combined, there emerges a new mechanism which may be called a habit-family hierarchy. The hypothesis is put forward that when an organism for any reason succeeds in solving a problem by a sequence of acts which is substantially that of one member of a habit family, the other members of the family, particularly those occupying a more favored position in the hierarchy, will automatically become active in the new situation without any specific practice whatever. This transfer may be thought of as being mediated mainly through the anticipatory reaction (rG) which is common to all members of the habit-family hierarchy and which therefore as a stimulus is conditioned to evoke the initial reactions of each member. It seems likely that this mechanism of transfer may prove to have an extremely wide application as an explanatory principle in many subtle and otherwise inexplicable forms of behavior at present usually designated indiscriminately as intelligence. The habit-family hierarchy is accordingly put forward as one of [p. 148] the basic mechanism of insight; presumably there are numerous other such mechanisms. Thus we seem to be on our way to an objective and discriminating definition of this immensely important but badly understood phenomenon.
In making the application of the principle to the particular problems of maze learning in the rat, it is assumed that a great number of habit families of the locomotor variety have been established by the locomotion of the animal in free space during its early life. From these principles a number of deductions may be made, though in some cases the certainty of the deductions is limited by uncertainty as to the facts concerning conditioned reflexes upon which they are based. The more important of these deductions are as follow:
1. Animals which through lack of practice or otherwise have not had a normal opportunity to set up initial habit-family hierarchies, should show little or no signs of transfer.
2. Other things equal, immediate transfer in the maze to a shorter path should be distinctly more effective where the drive stimulus in the old and the new situations is the same than where it is different.
3. In case an animal is specifically trained to go into a blind alley, he will, if the training has not been too extensive, eliminate the blind at the first opportunity to take the direct path.
4. If the training to take the blind has been very extensive, the animal will normally continue to take the blind if undisturbed, even after the door which would permit him to take the direct path is opened.
5. If, under these conditions, the animal is disturbed shortly before the choice point is reached, he will be likely to omit the blind and take the direct path.
6. Shortly after such a choice, provided the disturbing stimulus has ceased, the animal will be likely to revert to his accustomed use of the blind, though in general the tendency to take it will not be so strong as it was previously.
7. If such disturbing stimuli should repeatedly produce the choice of the shorter path, even though interspersed with a considerably larger number of choices involving entrances into [p. 149] the blind, the animal will come ultimately to choose the shorter path on most occasions.
8. The more frequently, relatively, choices are made as a result of disturbing stimulations, the more quickly will the animals acquire a fixed habit of choosing the direct path.
9. Animals in learning a maze will tend to show a predilection for blinds pointing in the direction of the food box, as contrasted with blinds originating near the same point and leading in the opposite direction.
10. Of two blind alleys pointing in the direction of the food box, other things equal, the one closer to it by direct line will be more favored by the animals and will be more difficult of elimination from the path.
11. Blinds pointing in the direction of the food box will show at first an increase in the percentage of entrances per trial, after which there will follow a gradual decrease. Blinds pointing away from the goal, other things equal, will show in general a progressive decrease in per cent of entrances from the beginning of practice.
12. On successive complete entrances to a blind pointing in the direction of the goal, the animal will proceed at a progressively slower pace especially as he approaches the blind end, whether the latter is visible or not.
13. The process of eliminating blinds pointing in the direction of the goal will be characterized by the animal turning around and coming out before reaching the end, and even before being able to see the blind end.
14. The depth of such partial entrances will tend to become less and less with continued practice, until finally the blind will not be entered at all.
15. The mean depth of all penetrations after the first partial entrance has taken place will average less for alleys pointing in the direction of the goal than for those pointing in the opposite direction.
16. Short blind alleys pointing away from the goal will be less readily eliminated by hungry animals than will long blinds.
17. Short blind alleys pointing in the direction of the goal [p. 150] I will be more readily eliminated by hungry animals than will long blinds.
18. After the animal has consistently eliminated a blind alley and has made repeated perfect runs, a slight disturbance just before he reaches the opening to such a blind will be likely to cause an entrance into the alley. A slightly stronger stimulus should cause an entrance also into alleys extending in the opposite direction. Such tendencies to enter blinds, however, should be of relatively short duration.
19. With distributed practice there should be a relatively greater difference between the ease of eliminating goalward and nongoalward-pointing blinds than there would be under immediately consecutive massed practice.
20. Alleys pointing in the direction of the food box should be eliminated with greater difficulty when the animal is under the influence of caffeine, whereas alleys pointing away should be eliminated with greater ease.
Several of the above deductions are known on the basis of experiment to be true, though many of them have never been subjected to experiment. The latter may, therefore, present an opportunity to test the truth of the basic hypothesis from which they logically flow. Hypotheses (in psychology too often called theories) are not matters of faith to be defended with religious zeal. On the contrary they are to be subjected to the most rigorous experimental tests possible at the earliest feasible opportunity. Those hypotheses which clearly fail by this test must be ruthlessly discarded. And, by the same token, no hypothesis which fails to yield such deductions (as distinguished from the intuitions of its originator or protagonists) has any status in science.
References
1. Dashiell, J. F., Direction orientation in maze running by the white rat, Comp. Psychol. Monog., 1930, 7, No. 32.
2. De Camp, J. E., Relative distance os a factor in the white rat's selection of a path. Psychobiol ., 1930, 20, 245-253.
3. Dennis, W., A study of learning in the white rat, J. Genet.. Psychol., 1930, 37, 294-308.
4. Ebbinghaus, H., Memory (Trans. by Ruger), New York: Teachers Coll., Columbia Univ., 1913 (original date, 1885).
5. Evans, C. L., Recent advances in physiology, 4th ed., Philadelphia: P. Blatkiston's Son and Co., 1930.
6. Gengerelli, J. A., Preliminary experiments on the causal factors in animal learning, J. Comp. Psychol., 1928, 8, 435-457.
7. Hall, M. E., Remote associative tendencies in serial learning, J. Exper. Psychol, 1928, ii, 65-67.
8. Higginson, G. D., Visual discrimination in the white rat, J. Exper. Psychol.,1926, 9, 331-347.
9. Hilgard, E. R., and Marquis, D. G., Acquisition, extinction, and retention of conditioned lid responses in dogs (to appear).
10. Hobhouse, L. T., Mind in evolution, London: Macmillan and Co. Ltd., 1926.
11. Holmes, S.J., The evolution of animal intelligence, New York: Henry Holt and Co., 1911.
12. Hull., C. L., Quantitative aspects of the evolution of concepts, Psychol. Monog .,1920, 28, No. 123.
13. ____., Simple trial-and-error learning: a study in psychological theory, Psychol. Rev ., 1930, 37, 241-256.
14. ____., Knowledge and purpose as habit mechanisms, Psychol. Rev., 1930, 37, 511-525.
15. ____., Goal attraction and directing ideas conceived as habit phenomena, Psychol. Rev ., 1931, 38, 487-506.
16. ____., The goal gradient hypothesis and maze learning, Psychol. Rev ., 1932, 39, 25-43.
17. ____., Differential habituation to internal stimuli in the albino rat, J. Comp. Psychol ., 1933, 16, 255-273.
18. Johnson, H. M., Audition and habit formation in the dog, Behav. Monog ., 1913-15, 2, No. 8.
19. Koffka, K., The growth of the mind (Trans. by Ogden), New York: Harcourt, Brace and Co. Inc.; London: Kegan, Paul, Trench, Trubner and Co. Ltd., 1925.
20. Lashley, K. S., The effects of strychnine and caffeine upon the rate of learning, Psychobiol., 1917, 1, 141-170.
21. Lepley W. M., A theory of serial learning and forgetting based upon conditioned reflex principles, Psychol. Rev., 1932, 39, 279-288.
22. Lumley, F. H., An investigation of the responses made in learning a multiple choice maze, Psychol. Monog., 1931, 42, No. 189.
23. ____, Anticipation as a factor in serial and maze learning, J. Exper. Psychol., 1932, 15, 331-342.
24. Maier, N. R. F., Reasoning in white rats, Comp. Psychol. Monog., 1929, 6, No. 29.
25. Pavlov, I. P., Conditioned reflexes, an investigation of the physiological activity of the cerebral cortex (Trans. by Anrep), London: Oxford Univ. Press, 1927.
26. Peterson, J., The effect of length of blind alleys on maze learning: an experiment on twenty-four white rats, Behav. Monog., 1916-19, 3, No. 15.
27. Robinson, E. W., and Wever, E. G., Visual distance perception in the rat, Univ. Calif. Publ. Psychol ., 1930, 4, 233-239.
28. Shipley, W. C., An apparent transfer of conditioning, J. General Psychol ., 1933,8, 382-39I.
29. Spence, K. W., The order of eliminating blinds in maze learning by the rat, J. Comp. Psychol ., 1932, 14, 9-27.
30. Spraggs. S. D. S., Anticipation as a factor in maze errors, J. Comp. Psychol., 1933, 15, 313-329
31. Switzer, S. A., Backward conditioning of the lid reflex, J. Exper. Psychol ., 1930, 13, 76-97.
32. ____., Disinhibition of the conditioned galvanic skin response. J. General Psychol., 1933, 9, 77-100.
33. Tolman, E. C., Purposive behavior in animals and men, New York and London: The Century Co., 1932.
34. Tolman, E. C., Sign-Gestalt or conditioned reflex? Psychol. Rev., 1933, 40, 246-255.
35. Tolman, E. C., and Honzik, C. H., 'Insight' in rats, Univ. Calif. Publ. Psychol., 1930, 4, 215-232.
36. Valentine, H. M., Visual perception in the white rat, J. Comp. Psychol ., 1928, 8, 369-375.
37. Wheeler, H. W., and Perkins, F. T., Principles of mental development, New York: Thomas Y. Cromwell Co., 1932.
38. Wolfle, H. M., Conditioning as a function of the interval between the conditioned and the original stimulus, J. General Psychol., 1932, 7, 80-103.
Footnotes
[14] To over-ride a principle or factor by opposing to it one or more factors which possess in the aggregate a greater strength should not be considered as a disproof of the existence and significance of the first principle. Thus, for a balloon or an airplane or a thrown object to rise from the earth in opposition to gravity is in no sense a proof of the non-existence of gravity or that gravity need not be taken into consideration, as many an aviator has found to his sorrow.
[15] Suppose that a given degree of perfection would be attained in ten repetitions. By (16), p. 32, each trial would increase the strength of the tendency to take the shortpath by .428 unit, whereas it would increase the strength of the tendency to take the long path by .096 unit. It follows that two runs on the short path would strengthen that tendency by .856, whereas eight runs on the long path would increase the strength of that tendency by only 8 X .096, or .768 unit. This would leave a net advantage of .088 to the tendency to take the short path.
[16] Both Dashiell (1) and Tolman and Honzik (33, p. 119 ff. ) have reported this significant phenomenon on the basis of experiment.
[17] On the principle of the goal gradient alone, assuming the latter to operate according to the logarithmic law and, in the interest of simplicity of exposition, assuming an approximately straight curve of learning, it would be possible for the choice finally to settle on path X , even though there should be distinctly more trials on Y than on X . Path X from point O is about 27 units long, and path Y , about 41. By Table 2 of (16), path X should have, at point O , an excitatory strength in the proportion of 1.81, whereas path Y would have a strength in the proportion of .77. Since .77 is contained in 1.81 twice with .27 left over, it follows that path X would finally come to preponderate over Y even if the former had initially only half as many choices of the latter.
[18] Several writers, perhaps the earliest of whom is Peterson (26), have noted the progressively shortening penetrations of animals into blinds. Perhaps the most convincing study illustrating this tendency is that by Robinson and Weaver (27), truly beautiful experiment. Though the conditions of this latter investigation different certain respects from those assumed in the above deduction, it is believed that the explanation here offered holds substantially for them also.
[19] It is easy to deduce from Lepley's hypothesis (21) that rote series of nonsense syllables will be more easily learned by distributed practice. Presumably the same principles apply to mazes, particularly it most of the alleys point away from the goal. This would explain the well-known economy in maze learning by distributed practice. The action of the Lepley principle and the present one are thus in opposite directions, with the evidence favoring the dominance of the former. Even so, the above hypothesis seems to demand that there should be relatively less advantage from distributed practice on mazes made up largely of goalwards pointing blinds than on mazes made up largely of blinds pointing away from the goal. At the very least the pre-dominant direction of the blinds becomes a matter of significance in maze studies involving distributed vs. massed practice.