Fig. 81.
Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
The Principles of Psychology
William James (1890)
CHAPTER XXIII. [*]
THE PRODUCTION OF MOVEMENT.
The reader will not have forgotten, in the jungle of purely inward processes and products through which the last chapters have borne him, that the final result of them all must be some form of bodily activity due to the escape of the central excitement through outgoing nerves. The whole neural organism, it will be remembered, is, physiologically considered, but a machine for converting stimuli into reactions; and the intellectual part of our life is knit up with but the middle or 'central' portion of the machine's operations. Let us now turn to consider the final or emergent operations, the bodily activities, and the forms of consciousness connected therewithal.
Every impression which impinges on the incoming nerves produces some discharge down the outgoing ones, whether we be aware of it or not. Using sweeping terms and ignoring exceptions, we might say that every possible feeling produces a movement, and that the movement is a movement of the entire organism, and of each and all its parts. What happens patently when an explosion or a flash of lightning startles us, or when we are tickled, happens latently with every sensation which we receive. The only reason why we do not feel the startle or tickle in the case of insignificant sensations is partly its very small amount, partly our obtuseness. Professor Bain many years ago gave the name of the Law of Diffusion to this phenomenon of general discharge, and expressed it thus: "According as an impression is accompanied with Feeling, the aroused currents diffuse themselves over the brain, leading to a general agitation of the moving organs, as well as affecting the viscera."
[p. 373] In cases where the feeling is strong the law is too familiar to require proof. As Prof. Bain says:
"Each of us knows in our own experience that a sudden shock of feeling is accompanied with movements of the body generally, and with other effects. When no emotion is present, we are quiescent; a slight feeling is accompanied with slight manifestations; a more intense shock has a more intense outburst. Every pleasure and every pain, and every mode of emotion, has a definite wave of effects, which our observation makes known to us; and we apply the knowledge to infer other men's feelings from their outward display. . . . The organs first and prominently affected, in the diffused wave of nervous influence, are the moving members, and of these, by preference, the features of the face (with the ears in animals), whose movements constitute the expression of the countenance. But the influence extends to all the parts of the moving system, voluntary and involuntary; while an important series of effects are produced on the glands and viscera -- the stomach, lungs, heart, kidneys, skin, together with the sexual and mammary organs. . . . The circumstance is seemingly universal, the proof of it does not require a citation of instances in detail; on the objectors is thrown the burden of adducing unequivocal exceptions to the law."[1]
There are probably no exceptions to the diffusion of every impression through the nerve-centres. The effect of the wave through the centres may, however, often be to interfere with processes, and to diminish tensions already existing there; and the outward consequences of such inhibitions may be the arrest of discharges from the inhibited regions and the checking of bodily activities already in process of occurrence. When this happens it probably is like the draining or siphoning of certain channels by currents flowing through others. When, in walking, we suddenly stand still because a sound, sight, smell, or thought catches our attention, something like this occurs. But there are cases of arrest of peripheral activity which depend, not on central inhibition, but on stimulation of centres which discharge outgoing currents of an inhibitory sort. Whenever we are startled, for example, our heart momentarily stops or slows its beating, and then palpitates with accelerated speed. The brief arrest is due to an outgoing current down the pneumogastric nerve. This nerve, when stimulated, stops or slows the heart-beats, and this [p. 374] particular effect of startling fails to occur if the nerve be cut.
In general, however, the stimulating effects of a sense-impression preponderate over the inhibiting effects, so that we may roughly say, as we began by saying, that the wave of discharge produces an activity in all parts of the body. The task of tracing out all the effects of any one incoming sensation has not yet been performed by physiologists. Recent years have, however, begun to enlarge our information; and although I must refer to special treatises for the full details, I can briefly string together here a number of separate observations which prove the truth of the law of diffusion.
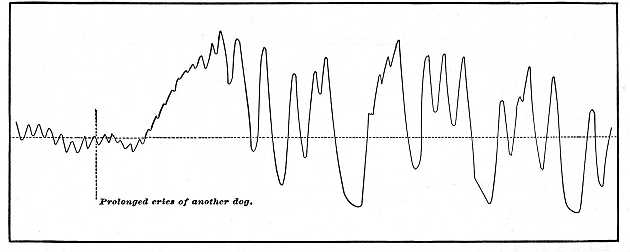
First take effects upon the circulation. Those upon the heart we have just seen. Haller long ago recorded that the blood from an open vein flowed out faster at the beat of a drum.[2] In Chapter III. (p. 98) we learned how instantaneously, according to Mosso, the circulation in the brain is altered by changes of sensation and of the course of thought. The effect of objects of fear, shame, and anger upon the blood-supply of the skin, especially the skin of the face, are too well known to need remark. Sensations of the higher senses produce, according to Couty and Charpentier, the most varied effects upon the pulse-rate and blood-pressure in dogs. Fig. 81, a pulse-tracing from these authors, shows the tumultuous effect on a dog's heart of hearing the screams of another dog. The changes of blood-pressure will occurred when the pneumogastric nerves were cut, showing the vaso-motor effect to be direct and not dependent on the heart. When Mosso invented that simple instrument, the plethysmograph, for recording the fluctuations in volume of the members of the body, what most astonished him, he says, "in the first experiments which he made in Italy, was the extreme unrest of the blood-vessels of the hand, which at every smallest emotion, whether during waking or sleep, changed their volume in surprising fashion."[3] Figure 82 (from Féré[4] ) [p. 375]
Fig. 81.
[p. 376] shows the way in which the pulse of one subject was modified by the exhibition of a red light lasting from the moment marked a to that marked b.

The effects upon respiration of sudden sensory stimuli are also too well known to need elaborate comment. We 'catch our breath' at every sudden sound. We 'hold our breath' whenever our attention and expectation are strongly
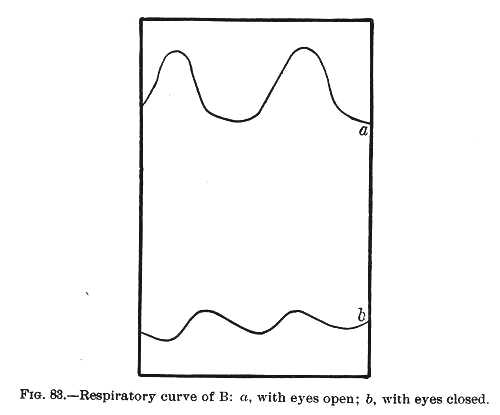
engaged, and we sigh when the tension of the situation is relieved. When a fearful object is before us we pant and cannot deeply inspire; when the object makes us angry it is, on the contrary, the act of expiration which is hard. I subjoin a couple of figures from Féré which explain them-[p. 377] selves. They show the effects of light upon the breathing of two of his hysteric patients.[5]

On the sweat-glands, similar consequences of sensorial stimuli are observed. Tarchanoff, testing the condition of the sweat-glands by the power of the skin to start a gal- [p. 378] vanic current through electrodes applied to its surface, found that "nearly every kind of nervous activity, from the simplest sensations and impressions, to voluntary motions and the highest forms of mental exertion, is accompanied by an increased activity in the glands of the skin."[6] On the pupil observations are recorded by Sanders which show that a transitory dilatation follows every sensorial stimulus applied during sleep, even if the stimulus be not strong enough to wake the subject up. At the moment of awakening there is a dilatation, even if strong light falls on the eye.[7] The pupil of children can easily be observed to dilate enormously under the influence of fear. It is said to dilate in pain and fatigue; and to contract, on the contrary, in rage.
As regards effects of the abdominal viscera, they unquestionably exist, but very few accurate observations have been made.[8]
The bladder, bowels, and uterus respond to sensations, even indifferent ones. Mosso and Pellicani, in their plethysmographic investigations on the bladder of dogs, found all sorts of sensorial stimuli to produce reflex contractions of this organ, independent of those of the abdominal walls. They call the bladder 'as good an æsthesiometer as the iris,' and refer to the not uncommon reflex effects of psychic stimuli in the human female upon this organ.[9] M. Féré has registered the contractions of the sphincter ani which even indifferent sensations will produce. In some pregnant women the fœtus is felt to move after almost every sensorial excitement received by the mother. The only natural explanation is that it is stimulated at such moments by reflex contractions of the womb.[10] That the glands are affected in emotion is patent enough in the case of the tears of grief, the dry mouth, moist skin, or diar- [p. 379] rhœa of fear, the biliary disturbances which sometimes follow upon rage, etc. The watering of the mouth at the sight of succulent food is well known. It is difficult to follow the smaller degrees of all these reflex changes, but it can hardly be doubted that they exist in some degree, even where they cease to be traceable, and that all our sensations have some visceral effects. The sneezing produced by sunshine, the roughening of the skin (gooseflesh) which certain strokings, contacts, and sounds, musical or non-musical, provoke, are facts of the same order as the shuddering and standing up of the hair in fear, only of less degree.
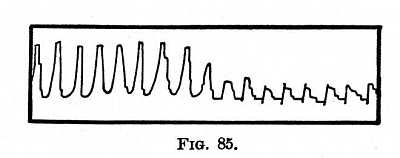
Effects on Voluntary Muscles. Every sensorial stimulus not only sends a special discharge into certain particular muscles dependent on the special nature of the stimulus in question -- some of these special discharges we have studied in Chapter XI, others we shall examine under the heads of Instinct and Emotion -- but it innervates the muscles generally. M. Féré has given very curious experimental proofs of this. The strength of contraction of the subject's hand was measured by a self-registering dynamometer. Ordinarily the maximum strength, under simple experimental conditions, remains the same from day to day. But if simultaneously with the contraction the subject received a sensorial impression, the contraction was sometimes weakened, but more often increased. This reinforcing effect has received the name of dynamogeny. The dynamogenic value of simple musical notes seems to be proportional to their loudness and height. Where the notes are compounded into sad strains, the muscular strength diminishes. If the strains are gay, it is increased. -- The dynamogenic value of colored lights varies with the color. In a subject[11] whose normal strength was expressed by 23, it became 24 when a blue light was [p. 380] thrown on the eyes, 28 for green, 30 for yellow, 35 for orange, and 42 for red. Red is thus the most exciting color. Among tastes, sweet has the lowest value, next comes salt, then bitter, and finally sour, though, as M. Féré remarks, such a sour as acetic excites the nerves of pain and smell as well as of taste. The stimulating effects of tobacco-smoke, alcohol, beef-extract (which is innutritious), etc., etc., may be partly due to a dynamogenic action of this sort. -- Of odors, that of musk seems to have a peculiar dynamogenic power. Fig. 85 is a copy of one of M. Féré's dynamographic tracings, which explains itself. The smaller contractions are those without stimulus; the stronger ones are due to the influence of red rays of light.
Everyone is familiar with the patellar reflex, or jerk upwards of the foot, which is produced by smartly tapping the tendon below the knee-pan when the leg hangs over the other knee. Drs. Weir Mitchell and Lombard have found that when other sensations come in simultaneously with the tap, the jerk is increased.[12] Heat, cold, pricking, itching, or faradic stimulation of the skin, sometimes strong optical impressions, music, all have this dynamogenic effect, which also results whenever voluntary movements are set up in other parts of the body, simultaneously with the tap.[13]
These 'dynamogenic' effects, in which one stimulation [p. 381] simply reinforces another already under way, must not be confounded with reflex acts properly so called, in which new activities are originated by the stimulus. All instinctive performances and manifestations of emotion are reflex acts. But underneath those of which we are conscious there seem to go on continually others smaller in amount, which probably in most persons might be called fluctuations of muscular tone, but which in certain neurotic subjects can be demonstrated ocularly. M. Féré figures some of them in the article to which I have already referred.[14]
Looking back over all these facts, it is hard to doubt the truth of the law of diffusion, even where verification is beyond reach. A process set up anywhere in the centres reverberates everywhere, and in some way or other affects the organism throughout, making its activities either greater or less. We are brought again to the assimilation which was expressed on a previous page of the nerve-central mass to a good conductor charged with electricity, of which the tension cannot be changed anywhere without changing it everywhere.
Herr Schneider has tried to show, by an ingenious and suggestive zoological review,[15] that all the special movements which highly evolved animals make are differentiated from the two originally simple movements, of contraction and expansion, in which the entire body of simple organisms takes part. The tendency to contract is the source of all the self-protective impulses and reactions which are later developed, including that of flight. The tendency to expand splits up, on the contrary, into the impulses and instincts of an aggressive kind, feeding, fighting, sexual intercourse, etc. Schneider's articles are well worth reading, if only for the careful observations on animals which they embody. I cite them here as a sort of evolutionary reason to add to the mechanical a priori reason why there ought to be the diffusive wave which our a posteriori instances have shown to exist.
I will now proceed to a detailed study of the more im- [p. 382] portant classes of movement consequent upon cerebromental change. They may be enumerated as --
1) Instinctive or Impulsive Performances;
2) Expressions of Emotion; and
3) Voluntary Deeds;
and each shall have a chapter to itself.
Footnotes
[*] Classics Editor's Note: The original mistakenly reads "Chapter XXII"
[1] Emotions and Will, pp. 4, 5.
[2] Cf. Féré. Sensation et Mouvement (1887), p. 56.
[3] La Paura (1884), p. 117. Compare Féré: Sensation et Movement, chap. XVII.
[4] Revue Philosophique, XXIV. 570.
[5] Revue Phil., XXIV. pp. 566-7. -- For further information about the relations between the brain and respiration, see Danilewsky's Essay in the Biologisches Centralblatt, II. 690.
[6] Quoted from the report of Tarchanoff's paper (in Plüger's Archiv, XLVI. 46) in the American Journal of Psych., II. 652.
[7] Archiv f. Psychiatrie, VII. 652; IX. 129.
[8] Sensation et Movement, 57-8.
[9] R. Accad. dei Lincei (1881-2). I follow the report in Hofmann u. Schwalbe's Jahresbericht, X. II. 93.
[10] Cf. Féré, Sensation et Movement, chap. XIV.
[11] The figures given are from an hysterical subject, and the differences are greater than normal. M. Féré considers that the unstable nervous system of the hysteric ('ces grenouilles de la psychologie') shows the law on a quantitatively exaggerated scale, without altering the qualitative relations. The effects remind us a little of the influence of sensations upon minimal sensations of other orders discovered by Urbantschitsch, and reported on page 29 of this volume.
[12] Mitchell in (Philadelphia) Medical News (Feb. 13 and 20, 1886); Lombard in American Journal of Psychology (Oct. 1887).
[13] Prof H. P. Bowditch has made the interesting discovery that if the reinforcing movement be as much as 0.4 of a second late, the reinforcement fails to occur, and is transformed into a positive inhibition of the knee-jerk for retardations of between 0.4' and 1.7'. The knee-jerk fails to be modified at all by voluntary movements made later than 1.7' after the patellar ligament is tapped (see Boston Med. and Surg. Journ., May 31, 1888).