An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3173
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
Posted March 2000
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE X
Irradiation and concentration of nervous processes in the cerebral hemispheres (continued): (b) Irradiation and concentration of inhibition over the entire cortex; (c) Irradiation and concentration of excitation.
In the last lecture it was shown how internal inhibition, initiated in a single definite point of the cortical part of an analyser, rapidly irradiates over the whole analyser, after which it is slowly concentrated upon its initial point. Furthermore, it was found that the progress of irradiation of internal inhibition can be traced step by step within the analyser.
In the present lecture we shall trace the progress of inhibition from one analyser to another over the whole cerebral cortex. Experiments bearing upon this process have been conducted for all the forms of internal inhibition recognized up to the present.
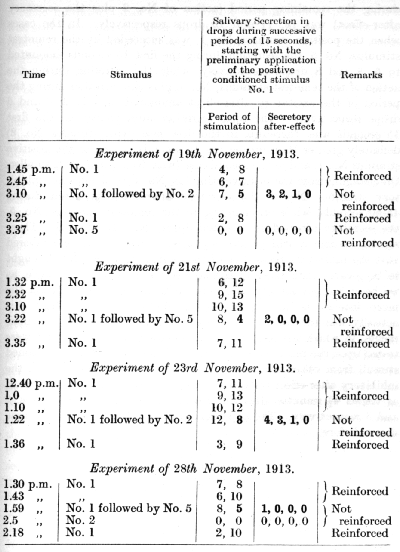
It was mentioned in the description of differential inhibition that when the inhibition is initiated in one analyser it reveals itself in other analysers as well, in the form of an inhibitory after-effect. In a series of experiments by Dr. Beliakov it was found that differential inhibition of a small intensity does not involve in its after-effects any other analyser than the one in which the inhibition was primarily developed, but that when the intensity of the inhibition is great the inhibitory after-effect involves other analysers as well. However, secondary inhibition of other analysers is in every case much weaker than that of the- analyser in which the differentiation was primarily developed. A number of experiments will be described to illustrate these relations:
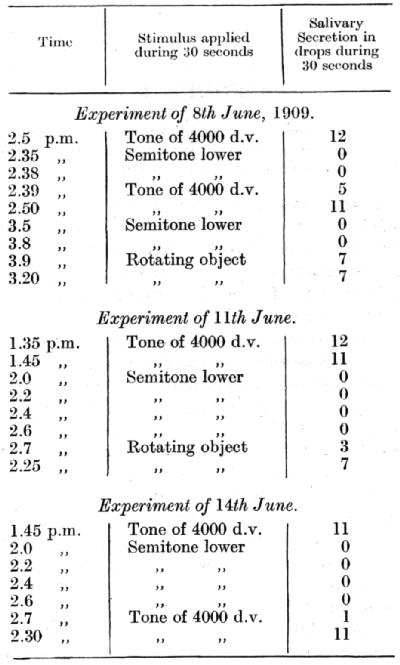
A conditioned alimentary reflex was established to a tone of 4,000 d.v. produced by a Galton's whistle; from this a pitch of one semitone lower was differentiated. A second positive conditioned alimentary reflex was established to a noiselessly rotating object. The reflex to the auditory stimulus gave an average secretion of 11-12 drops during 30 seconds, and the reflex to the visual stimulus gave an average of 7-8 drops. The results of a series of experiments [p. 168] upon the irradiation of differential inhibition are illustrated by the following table :
The significance of these experiments is obvious. After the inhibitory semitone had been repeated twice with an interval of 3 minutes between the successive repetitions, the positive tone tested half a minute later had lost more than 50% of its effect, while the visual stimulus showed no diminution in its effect at all (experiment of 8th June). After a greater summation of the inhibitory after-effect by four successive repetitions of the semitone with intervals of 2 minutes between them, the reflex to the visual stimulus [p. 169] test half a minute later (in the experiment of 11th June) showed a stimulus (tested in the experiment of 14th June) was nearly complete.
Similar results are obtained with extinctive inhibition, as shown in a series of experiments by Dr. Gorn. In these experiments, owing to the introduction of certain modifications into the technique, the irradiation of the inhibitory process was even more clearly exhibited, especially as regards the earlier removal of its after-effect from the secondarily involved analysers. The following are some of the experiments:
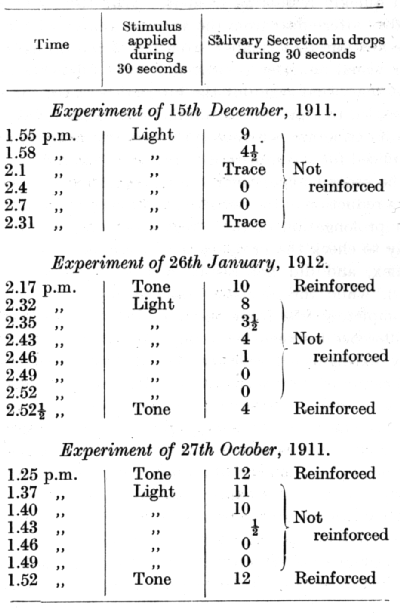
Separate conditioned alimentary reflexes were established in a dog to the tone c' sharp of a pneumatic tuning-fork and to flashes of three electric lamps, each of 16 candle-power.
 [p. 170]
[p. 170]
The conditioned reflex to light was in these experiments subjected to extinction, and the inhibitory after-effect was tested upon the reflex to the auditory stimulus. The reflex to the visual stimulus itself, after extinction up to the point of two consecutive zeros, showed the first signs of spontaneous recovery only after an interval of 231/2 minutes (15th December, 2.31 p.m.). The reflex to the auditory stimulus, on the other hand, when tested immediately after a similar extinction of the visual reflex was diminished in strength by only 60% (26th January, 1912, 2.521/2 p.m.), and when tested in the third experiment after a pause of 21/2 minutes it was quite free of any inhibitory after-effect (27th October, 1911, 1.52 p.m.). It is thus seen that the inhibition irradiated from the visual to the acoustic analyser-never, however, producing a full inhibitory after-effect, and quickly retreating from it. That the rapid removal of the inhibitory after-effect from the acoustic analyser in these experiments was not due to a smaller intensity of the primary inhibition within the visual analyser is proved by the following details. In the case of the visual stimulus the interval between the repetitions (21/2 minutes) led to a progressive diminution of the reflex through all the stages of extinction, whereas in the acoustic analyser an identical interval sufficed for a complete restoration of the reflex. Moreover, when in the second experiment the extinction of the visual reflex had led to a reduction of the secretion to 31/2 drops as against the usual 8 drops, a prolongation of the interval on purpose to 71/2 minutes served only to check the progress of the diminution in the strength of the reflex, and did not result in any appreciable restoration (2.43 p.m.), while the acoustic analyser was after that interval already completely free from any inhibitory after-effect. Neither was the difference between the restoration of the primarily and the secondarily extinguished reflexes due to any peculiarities of the cortical elements of the visual and auditory cells, as was proved by experiments in which the tone was subjected to the primary extinction, and the effect of secondary inhibition was observed on the visual analyser. The relations between the rates of recovery from the inhibition in the primarily and secondarily extinguished analysers in these experiments were exactly the same as before, and there is no need, therefore, to give separate tables. It can now reasonably be assumed that in the secondarily extinguished analyser we deal with the periphery of the irradiated inhibition, and that its recessional movement begins at the periphery and proceeds towards the point [p. 171] of initiation of the inhibition exactly in the same manner ass within the single cutaneous analyser (concentration of inhibition).
Experiments on irradiation of extinctive inhibition over the entire cortex of the hemispheres were performed with a great number of variations. For example, two conditioned reflexes were developed in the analyser which was subjected to primary extinction (e.g. visual analyser), and the inhibitory effect of experimental extinction of one of these reflexes was observed upon both of them and also upon reflexes belonging to other analysers. Recovery from the inhibitory after-effect was found first of all in secondarily extinguished reflexes belonging to other analysers. Shortly afterwards recovery occurred in the secondarily extinguished reflex belonging to the visual analyser itself, and finally, but after a much longer time, the reflex which underwent the primary extinction became freed from the inhibitory after-effect. The difference in the length of time necessary for the recovery of the two reflexes belonging to the primarily inhibited visual analyser clearly points to a regional localization of reflexes within the analysers, such as was already suggested in the preceding lecture.
Experiments performed with irradiation of conditioned inhibition gave, on the whole, very similar results. One definite additional stimulus was used as a common conditioned inhibitor for conditioned reflexes belonging to different analysers, each of the conditioned stimuli in turn being repeated with the additional stimulus and remaining in the compound always without reinforcement, so that the compound conditioned stimulus developed inhibitory properties. The actual experiments were conducted as follows. First one of the conditioned stimuli was applied, and its secretory effect recorded. Next the conditioned inhibitor was applied in combination with the same or another conditioned stimulus, and the combination was repeated several times in succession. Finally, the effect of the first conditioned stimulus applied singly was again tested at different intervals of time after the last application of the inhibitory combination. When the positive reflex tested was the one employed in the inhibitory combination, it was found to be considerably inhibited, and was only slowly freed from the inhibitory after-effect; when, however, the inhibitory combination was made with a stimulus from another analyser, the stimulus undergoing the test was only slightly inhibited, and rapidly recovered its original strength. It is thus seen that the inhibitory after-effect was very powerful and prolonged [p. 172] in the analyser of the reflex subjected to the primary conditioned inhibition, but was only weak and of short duration in other analysers.
In a series of experiments conducted by Dr. Degtiareva separate conditioned alimentary reflexes were established to metronome (M) and to flashes of electric lamps (L). A noiselessly rotating object (R) was used as a conditioned inhibitor common to both positive stimuli. The application of the conditioned stimuli both singly and in the inhibitory combination was always continued during one minute, and the amount of saliva secreted during this time was recorded. The following experiments give the final result of the determination Of the minimal interval which was necessary for complete recovery of the conditioned reflexes from their secondary inhibition.

These experiments show that after five repetitions of the inhibitory combination of which the metronome formed part, the minimum period necessary for the complete recovery of the positive reflex to the metronome was 7 minutes (experiment of 13th May, 4.46 p.m.). On the other hand, the minimum period required for the complete restoration of the reflex to the metronome after five repetitions of [p. 173] the inhibitory combination of rotating object plus visual stimulus was only 15 seconds (experiment of 16th May, 4.351/4 p.m.).
Finally, we have studied the irradiation of that form of internal inhibition which develops during the first phase of reflexes with a long delay. Owing to the peculiar character of this type of internal inhibition, occurring as part of a diphasic phenomenon, it might naturally be expected to exhibit peculiarities with regard to its after-effect, and such was actually found to be the case. The results obtained in experiments with different dogs varied considerably, and only in a certain number of cases could the irradiation of the inhibition easily be followed over the entire cortex. The experiments were performed in the following manner. The stimulus for the delayed reflex was continued only during the first part of the inhibitory phase of the delayed reflex, and was then followed either immediately or after some interval of time by a conditioned stimulus from another analyser ; the strength of this stimulus had previously been determined, and was now again tested to find the magnitude of the inhibitory after-effect. Some of these experiments are given below.
Two conditioned reflexes delayed for 30 seconds were established, the conditioned stimulus being in the first case flashes of electric lamps, and in the second tactile stimulation of the skin; a third conditioned reflex to a metronome was delayed for 3 minutes; the inhibitory phase of this reflex was equal to 11/2-2 minutes: all three reflexes were established with the help of acid.
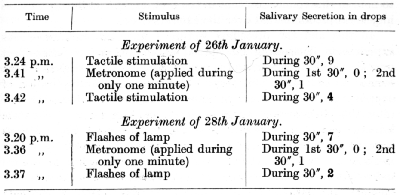
The result is quite definite. The inhibition evoked by the metronome in the first stage of its action has irradiated from the [p. 174] acoustic analyser into both the visual and tactile cutaneous analysers, diminishing their reflexes by 60-70%.
In another dog the results were almost completely reversed. The conditioned stimuli applied immediately after the inhibitory phase of the delayed reflex produced an increased positive effect. The reason of this difference Was found without difficulty. In the first dog, when an interval of 2 minutes was introduced between the isolated action of the metronome and the application of the succeeding conditioned stimulus, the excitation phase of the delayed reflex never appeared during these 2 minutes. In other words, the inhibitory phase of the delayed reflex in this dog was effectively isolated from the excitatory phase. On this account, under the conditions of the experiments, the inhibition became obvious in its after-effect upon both the visual and tactile Stimuli. In the second dog, on the other hand, when. the same interval was introduced between the isolated action of the metronome and the application of the succeeding conditioned stimulus, the active phase of the reflex to the metronome became revealed during the interval. In this dog, therefore, under the conditions of the experiment, the effect of the positive conditioned stimuli falling within the positive phase of the delayed reflex was not diminished but considerably increased on account of summation of the two reflexes. In this second dog the excitatory process appeared earlier and was more powerful; in other words, the separation of the inhibitory phase of the delayed reflex from the excitatory phase was not complete. It is interesting to note in this connection that in the first dog the inhibitory process in general predominated over the excitatory one.
In this and the preceding lecture I have adduced numerous experiments illustrating the spread of the inhibitory after-effect over the cerebral cortex, and it may be that to some readers I have seemed to over-stress this point. However, such multiplication of experiments was necessary in order to emphasize sufficiently how often this phenomenon occurs in simple and pure forms. The facts hitherto considered present only the fundamental interrelations between the inhibitory and excitatory processes revealed by special methods of experimentation, and often merely owing to special individual peculiarities of the animal. In many cases these fundamental interrelations are obscured by various accessory elements which render the whole process much more complex.
At the very beginning of our investigation of the irradiation of [p. 175] internal inhibition we were struck by the paradoxical and unexpected results of certain experiments [Dr. Krasnogorsky]. Dr. Krasnogorsky's experiments on the irradiation and concentration of differential inhibition within the cutaneous analyser were described in the preceding lecture (page 153). The preliminary test of a conditioned reflex to tactile stimulation of thc skin at a place situated at a distance of 22 cm. from thc place primarily inhibited elicited s drops of salivary secretion during 30 seconds, while a visual reflex gave a secretion of 5 drops. One minute after the last of three successive stimulations of the inhibitory place the visual reflex was found to be reduced to zero; in other words, the inhibitory after-effect spreading from the tactile into the visual analyser had completely overshadowed the excitatory effect of the visual stimulus. After ten minutes' interval the inhibitory place was again stimulated, now four times in succession, and on testing the tactile reflex at the place 22 cms. away, one minute after the last inhibitory stimulus, it was found to be completely free from inhibition.
Similar phenomena, have been recorded in experiments on conditioned inhibition conducted by Dr. Chebotareva. Several conditioned reflexes were established in a dog to stimuli belonging to different analysers. A conditioned reflex to a metronome, when applied in conjunction with a conditioned inhibitor in the form of a visual stimulus, failed to elicit a single drop of saliva. A minute or two later the metronome applied singly (when, of course, it was reinforced) produced its full secretory effect. Conditioned reflexes to a tactile stimulus and to an odour of camphor, however, when tested shortly afterwards were still found to be considerably inhibited.
It appears, therefore, from these experiments that, contrary to our expectation, the reflexes recovered from the inhibitory after-effect in the primarily inhibited analyser sooner than in any other. However, on careful consideration of the records of the development of the various reflexes in the animals, it became obvious that these apparently anomalous results were obtained only in cases when the secondarily inhibited reflexes belonging to other analysers were but recently established or, in the case of old reflexes, when they had not been exercised for a considerable time or were weak through having been established with the help of stimuli of small intensity. Hence it follows that if a certain area which was originally under the influence of inhibition has become free from the inhibitory after-effect, [p. 176] it does not necessarily mean that the inhibition is also withdrawn from the regions surrounding this area: under certain definite conditions thc process of excitation in some areas of the cortex may prevail over the co-existing and somewhat weakened inhibition of the surrounding areas. It is, indeed, our experience that conditioned reflexes in which the phase of initial generalization has not been succeeded by sufficient specialization, as well as reflexes of small intensity of excitation, become very easily disturbed, both by external and by internal inhibition. Those strong conditioned reflexes in which the specialization has been carried to a high degree, on the other hand, are not so easily influenced by external or internal inhibition. The question of stability and of resistance of strong and well-developed positive and negative conditioned reflexes will be considered in the thirteenth lecture.
In order to elucidate these exceptional phenomena, which appeared to contradict the. more general rule for the irradiation and concentration of inhibition, the following experiments were specially conducted by Dr. Pavlova.
Conditioned reflexes were established to several stimuli belonging to different analysers, particular attention being directed to maintaining an equal number of repetitions of all the various stimuli, always, of course, with their reinforcement. After the maximum intensity of every one of the reflexes had been reached, a conditioned inhibition was developed for one of the reflexes by the usual method of non-reinforcement of the corresponding stimulus when applied in combination with the conditioned inhibitor. The strengthening of the conditioned inhibition was carried to the point when the inhibitory after-effect as tested by the application of the corresponding stimulus became limited to two minutes. Of course, during the period of development of the conditioned inhibition all the positive reflexes continued to be reinforced an equal number of times. At this stage of the experiment all the other stimuli, which up till then had not been combined with the conditioned inhibitor, were now for the first time tested in company with the inhibitor. It was found that these reflexes also, with only one exception, became free of inhibitory after-effect two minutes after initiation of the conditioned inhibition. The exception in question was a weak conditioned reflex to the appearance -of a somewhat dim light, which was found to be still inhibited for some time after the recovery of the others. From these experiments it becomes clear [p. 177] that one and the same intensity of inhibition, when it had ceased to produce any diminution of the stronger reflexes, nevertheless exercised a definite action upon the weaker ones.
This dependence of the inhibitory after-effect upon the intensity of the secondarily inhibited reflexes is only one of several factors complicating the otherwise smooth progress of irradiation and concentration of inhibition. In the next lecture we shall describe another factor of much greater importance, producing a more profound disturbance in the progress of irradiation and concentration.
We shall turn now to a discussion of the experimental evidence with regard to the after-effect of the excitatory process; this has not, however, yet been studied to so great an extent as the after-effect of the inhibitory process.
The first experiments upon the after-effect of excitation were conducted by Dr. M. Petrova on similar lines to the early experiments on the irradiation and concentration of inhibition. Five separate apparatuses for tactile stimulation of the skin were arranged at equal distances apart along the hind limb of a dog, the first of the series being placed over the paw and the fifth over the pelvis. In one animal the tactile stimulation of the paw (No. 1) served to evoke a conditioned reflex to acid, while in another animal it served to evoke a conditioned alimentary reflex. Owing to initial generalization of the reflex, the stimulation of the remaining places (Nos. 2, 3, 4 and 5) also evoked at first positive alimentary reflexes or defence reflexes to acid. These places, however, rapidly became differentiated by the method of contrast, giving finally a zero effect, so that only the stimulation of the paw remained positive. At the beginning of each experiment the tactile stimulation of the paw (the positive stimulus) was applied during 30 seconds, and the amount of salivary secretion was recorded during the first half and during the second half of the period of stimulation. After 30 seconds of stimulation the reflex was reinforced. Then, after some interval of time, the paw was again stimulated, but now the stimulation was continued during only the first 15 seconds, and was followed immediately by stimulation of one or another of the inhibitory places during the second 15 seconds. Results were obtained as follows. On changing the stimulus from the positive place (No. 1) to the nearest inhibitory place (No. 2) the salivary secretion during the second [p. 178] period of 15 seconds was diminished only slightly, or even not at all, as compared with the secretion observed during the first period of 15 seconds (action of apparatus No. 1). But when the remotest apparatus (No. 5) acted during the second period of 15 seconds, the salivary secretion was considerably diminished. Stimulation of the places Nos. 3 and 4 under identical conditions gave rise to a secretion intermediate between that evoked at places Nos. 2 and 5, the secretory effect at place No. 3 exceeding that of place No. 4.
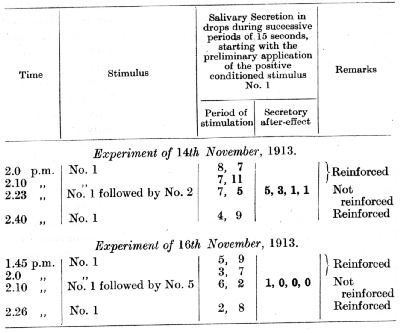
In further experiments not only was the salivary secretion during the 15 seconds of the actual stimulation of the inhibitory places recorded, but also the entire secretory after-effect. In these experiments the difference in the effect of the nearer and the more remote stimuli was exhibited almost without exception and with great distinctness. For example, stimulation of the nearest place (No. 2) evoked a far more prolonged and greater secretion than did stimulation of the remotest place (No. 5). The following experiments illustrate this case. The dog used in these experiments had its tactile conditioned reflexes established with the use of acid.
 [p. 179]
[p. 179]
The results of these experiments are very definite. In the cases when the positive stimulus, No. 1, was succeeded by the nearest inhibitory one, No. 2, the amount of salivary secretion recorded during the first 15 seconds (action of No. 1) was in the three different experiments 7, 7, 12 drops respectively, while the secretion [p. 180] during the remaining period (action of No. 2 plus the secretory after-effect) was 15, 11 and 16 drops respectively. In the cases when the positive stimulus, No. 1, was succeeded by the remotest stimulus, No. 5, the secretion during the first 15 seconds amounted to 6, 8 and 8 drops respectively, while the secretion during the action of the remotest stimulus, No. 5, plus the secretion during the period of the excitatory after-effect amounted to only 3, 6 and 6 drops respectively. The total secretion subsequently to the first 15 seconds was, therefore, in the first case (with stimulus No. 2) definitely greater than during the 15 seconds' action of the positive stimulus No. 1, and in the second case (with No. 5) definitely smaller. On changing from the positive stimulus, No. 1, to the nearest inhibitory stimulus, No. 2, the secretory after-effect persisted during 45-60 seconds, while on changing to the remotest stimulus, No. 5, the secretory after-effect persisted during only 15 seconds, These results assume a still greater significance when it is remembered that the inhibitory properties of the nearest stimulus, No. 2, ought to be greater than those of the remotest stimulus, No. 5, since, as we have learned already, the more delicate the differentiation the more intense is the underlying inhibitory process. That this actually was the case was definitely established by special experiments, in which the inhibitory after-effects from Nos. 2 and 5 were tested upon the positive reflex from No. 1 and upon some conditioned stimuli from other analysers. The following is an example of the inhibitory after-effect upon the positive reflex from the place No. 1, as tested 41/2 minutes after the use of the inhibitory stimuli Nos. 2 and 5 respectively. The experiments were carried out on the same dog as before.
 [p. 181]
[p. 181]
The inhibitory after-effect following stimulation of place No. 2 was still very powerful after an interval of 41/2 minutes, whereas the inhibitory after-effect following stimulation of place No. 5 was negligible.
In view of the results of these further experiments, there seems to be no reason to doubt that the excitation evoked by the positive stimulus in the previous experiments exerted a greater effect on the place nearest to it then on the place most remote. The whole phenomenon must, therefore, be regarded as a result of an irradiation of excitation.
Some experiments upon the irradiation of excitation have recently been repeated by Dr. Podkopaev. The places for tactile stimulation were taken on the skin of one side of the body, the first apparatus being arranged over the fore paw and the remaining apparatuses being placed at equal distances apart along the trunk and hind limb of the dog, the last being arranged over the hind paw. Stimulation of the area over the fore paw was given the properties of a positive alimentary conditioned stimulus, while stimulations of the remaining areas were given the properties of negative stimuli. The salivary secretion was recorded during successive periods of five seconds. Usually the positive conditioned stimulus was continued during 30 seconds and was then immediately reinforced, but for the purpose of the tests the stimulus was occasionally allowed to act during only 15 seconds, and was followed by a pause of 15 seconds before the reinforcement. In this manner it was possible to follow the course of the secretion during 30 seconds when the stimulus was acting throughout, and also during 30 seconds when the stimulus was abbreviated to the first 15 seconds. With the longer stimulation the ratio of the secretion during the first 15 seconds to that during the second 15 seconds came out as 1:2.4; in the case of the abbreviated stimulation it came out as 1:1.25. In the next stage of the experiments the positive stimulus after acting during the first 15 seconds was followed immediately by an inhibitory stimulus which acted during the succeeding period of 15 seconds, and this stimulus was given by stimulation of a nearer or a remoter place. The ratio of the amount of secretion during the first 15 seconds to that during the second 15 seconds came out in the former case as 1:1.35. and in the latter case as 1:0.53. It is obvious, therefore, that in this case, as in the previous experiments, the excitatory after-effect of the positive stimulus [p. 182] was strong in places nearest to it and negligible in the remoter places.
Neither this last nor the preceding series of experiments permits us to determine with which phase of the movement of excitation we have to deal, whether the phase of irradiation or the phase of concentration. Experiments are at present being conducted in which the inhibitory stimuli are applied, not immediately on cessation of the positive stimulus, but after a short pause of varying length. Unfortunately, the direct experimental data concerning irradiation and concentration of excitation at our disposal are limited to the facts described above. There are, however, a number of isolated observations which have some bearing upon this subject, and these will now be described.
One of our animals -- a sheep-dog -- had a very exaggerated form of what might be called "the warding reflex." The experimenter [Dr. Besbokaya], who always worked with the dog in a separate room, used to prepare it for the experiment without the slightest difficulty, placing it in the stand and adjusting the various apparatuses for recording and for stimulation. Under Dr. Besbokaya's control the experiments on conditioned reflexes proceeded quite smoothly, but the moment a stranger appeared in the experimenting room the dog always developed a powerful aggressive reaction, especially if the stranger (in that case myself) took the place of the experimenter. Applying the established conditioned alimentary stimulus under these circumstances, I always obtained a secretory reflex considerably above normal, the dog consuming its food with obviously exaggerated muscular effort. This aggressive reaction evoked by my presence became diminished in intensity when I intentionally kept quite still: the only part of the reflex still remaining was the gaze of the animal, which was kept fixed on me. The conditioned alimentary stimulus now produced a much smaller salivary secretion than the normal, and sometimes even no secretion at all. As soon, however, as I relaxed my attitude, and especially if I moved about, the aggressive reaction returned in full vigour, and the salivary and motor reactions in response to the conditioned alimentary stimulus became considerably increased in their intensity. It is interesting to note that although this dog grew calm in the presence of a stranger who kept quite still, the aggressive reaction returned after each reinforcement after the end of the conditioned stimulus, the reaction continuing for some time, [p. 183] even though the stranger remained motionless. There appears to be only one possible explanation of this phenomenon. The powerful excitatory effect of the intruder and his movements must have irradiated over the hemispheres, and increased amongst others the excitability of that area which is related to alimentary functions. When, however, this excitation diminished on account of weakening and concentrating of the external stimulus the reverse took place, and the excitability of remoter areas became diminished as compared with the normal (effect of external inhibition). In a similar manner the eating of food temporarily heightened the excitability of the centre for the aggressive reaction.
A similar phenomenon was observed in experiments conducted [by Dr. Prorokov] with another exceptional dog. In this dog tactile stimulation of the skin evoked a peculiar and very definite motor reaction, akin to a reaction to tickling, and possibly of a sexual character. It elicited also a conditioned alimentary reflex which was abnormally powerful, exceeding any of the other conditioned alimentary reflexes to non-tactile stimuli. As a rule, tactile stimulation of the skin evoked first of all its own special motor reaction, and was then succeeded after 10-l5 seconds by an exceedingly powerful alimentary motor reflex. When the special motor reaction was overcome by the use of a technique which will be discussed in the last lecture, the conditioned alimentary reflex to the tactile stimulus became reduced to a normal level, and became less powerful, for example, than homogeneous reflexes to auditory stimuli. This phenomenon must unquestionably be regarded as a further example of irradiation of excitation.
We are inclined to give a similar interpretation also to the following fact. In the case of integration of conditioned reflexes based upon the use of different alimentary substances, the inhibition of some of them by others does not develop immediately, but only after an interval of some minutes. It is quite conceivable that at first an irradiation of excitation is initiated from the cortical point corresponding to some definite alimentary substance. The excitability of neighbouring points thereby becomes temporarily increased, and it is only afterwards, when the excitation gets weakened and concentrated, that the inhibition reveals itself [experiments of Dr. Savitch].
A similar argument can be advanced in explanation of some unexpected results obtained by Dr. Vassiliev in the course of formation [p. 184] of heterogeneous conditioned reflexes to thermal stimuli. A conditioned reflex to acid was established by the application of a thermal stimulus of 0°C. to the skin of the animal, and a conditioned alimentary reflex by means of a thermal stimulus of 47°C. The development of these reflexes required a considerable time, and when finally established they were found to show the following extremely interesting peculiarity. If the conditioned reflex to cold (based on acid) was repeated with reinforcement several times in succession, the alimentary conditioned reflex to heat when evoked directly afterwards became transformed into a defence reflex to acid similar to that obtained with the stimulus of cold. Reversely, cold applied after several reinforcements of the conditioned alimentary reflex to heat brought about an alimentary reaction instead of the customary reaction to acid. The fact of this transformation was quite distinct and beyond doubt, since the nature of the reflex could easily be established by, for example, observing the motor reactions of the animal, which differ considerably for the two reflexes. In the case of the alimentary reflex the dog turned towards the experimenter, dividing its gaze between him and the source of food and smacking its lips; in the case of the reflex to acid the dog turned away from the experimenter, snorting and whining, shaking its head and making ejective movements with its tongue. Moreover, the reflexes could easily be distinguished by the composition of the saliva from the submaxillary salivary gland, which in the case of the alimentary reflex was thick and mucous, and in the case of the reflex to acid, fluid or watery. These points of difference were established in the given case with especial care. The persistent and prolonged transformation of one reflex into the other was the more strange and unexpected, since such a confusion of heterogeneous reflexes never occurs in other analysers; for example, in the acoustic analyser, even if the two stimuli to heterogeneous reflexes differ by less than a tone no transformation occurs, although it might be thought that the difference between two neighbouring tones was of much smaller vital significance than the difference between heat and cold.
The explanation turns on a certain peculiarity in the relations of the analysers for heat and cold. As is well known the peripheral ends of these analysers are interspersed with one another over the surface of the skin. Now it has been proved experimentally by Dr. Shishlo that the cortical ends of the two analysers overlap and if is [p. 185] tempting, therefore, to conjecture that the cortical ends are interspersed similarly to the peripheral ends. In that case irradiation from the one analyser to the other would be facilitated and such confusion between the reflexes as is illustrated in the above experiments can be accounted for.
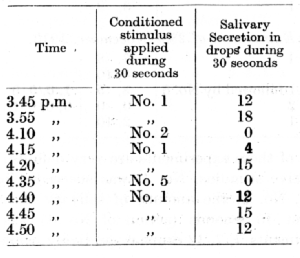
I shall now return to a further consideration of a phenomenon which was referred to in the preceding lecture in connection with the analysing activity of the cortex, but which, from the point of view of its intimate mechanism, has to be dealt with in this section. This concerns the initial generalization of any recently established conditioned stimulus, representing an apparently spontaneous positive activity of stimuli which had never previously been applied, but which constitute an allied group with the stimulus which had acquired conditioned properties on account of reinforcement. As an example we may take a series of experiments on the cutaneous analyser conducted by Dr. Anrep. Six apparatuses for tactile stimulation of the skin were arranged at definite places along one side of the body. The action of apparatus No. 0 at a place on the thigh was given positive alimentary conditioned properties in a short trace reflex. The remaining places were distributed as follows :
No. 1 on the hind paw, No. 2 on the pelvis, No. 3 on the middle of the trunk, No. 4 on the shoulder, No. 5 on the fore leg, No. 6 on the front paw. Stimulation of any of the latter places produced a spontaneous conditioned secretory effect, although none of them had been combined with food; the intensity of their secretory effect decreased very gradually with the distance from the place on the thigh, No. 0. The table gives the average figures obtained with one of the three dogs employed for these experiments. The amount of saliva is expressed in drops, each equal to 0.01 c.c. The secretion was recorded for the 30 seconds during which the stimulation lasted.
![]()
The stability of generalization varies considerably in different dogs, and it is not always easy to keep it on a definite level during a series of experiments. If the stimuli were applied to new places, and reinforced every time, the reflexes would become completely generalized. On the other hand, if these stimuli remained without [p. 186] reinforcement an opportunity would be provided for the development of a differential inhibition. In view of this, and in order to secure more or less exact and comparable results which could be verified by repetition, the stimuli were alternately applied with and without reinforcement, or else the applications were repeated at long intervals of time (2-3 weeks).
It remains now to inquire into the intimate mechanism of generalization, and the following interpretation seems best to agree with our present knowledge. It may be assumed that each element of the receptor apparatus gains representation is the cortex of the hemispheres through its own proper central neurone, and the peripheral grouping of the receptor organs may be regarded as projecting itself in a definite grouping of nervous elements in the cortex. A nervous impulse reaching the cortex from a definite point of the peripheral receptor does not give rise to an excitation which is limited within the corresponding cortical element, but the excitation irradiates from its point of origin over the cortex, diminishing in intensity the further if spreads from the centre of excitation, exactly as we have seen in the case of inhibition. Just as the initial point of cortical excitation becomes connected with the centre of the unconditioned reflex, so also do the secondary points of cortical excitation, and this leads to the formation of many accessory reflexes. These reflexes decrease progressively in strength with increasing distance of the secondarily excited areas from the point of primary cortical excitation, since the magnitude of the conditioned reflex is rigidly determined by the intensity of excitation. This interpretation agrees with Dr. Koupalov's observation that an experimental generalization of a localized tactile reflex develops gradually and progressively, and that complete equalization of the effect of stimulating any place of the skin is only reached after a considerable time. This, of course, is only natural, since conditioned reflexes from points further removed from the primary excitation appear considerably later, the accessory tactile reflexes being much weaker to start with at the remoter places
The explanation advanced as regards the initial generalization of simultaneous conditioned reflexes within a single analyser can be effectively applied to the type of universal generalization which is observed in long-trace reflexes, (Lecture VII). In the case of the long pause between the conditioned and the unconditioned stimulus, the excitation initiated by the conditioned stimulus has time to [p. 187] spread over the entire cortex, and in this manner all the individual cortical points can now link up with the centre for the unconditioned reflex as soon as this becomes excited. In the case of s short pause between the two stimuli the centre for the unconditioned reflex determines a concentration of the conditioned excitation towards itself, thus limiting any extensive irradiation over the general mass of the hemispheres.