
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3173
(Return to index)
By Ivan P. Pavlov (1927)
Translated by G. V. Anrep (1927)
Posted December 2000
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE XIII
The cortex as a mosaic of functions: (a) Examples of the mosaic character of the cortex and the more obvious ways in which this character is acquired. (b) Variability of the physiological properties of different points of the cortex in some instances, stability in others. -- The cortex as a united complex dynamic system.
It becomes obvious from all the previous discussion that through The medium of the cerebral cortex a great number of environmental Changes establish now positive, now negative, conditioned reflexes, And determine in this manner the different effector activities of the animal organism and its everyday behaviour. All these conditioned reflexes must have definite representation in the cerebral cortex in one or another definite group of cells. One such group of cells must be connected with one definite activity of the organism, another group with another activity: one group may determine a positive while another may inhibit an activity. The cerebral cortex can accordingly be represented as an exceedingly rich mosaic, or as an extreme complexity as a switchboard." However, in spite of its extreme complexity as a switchboard, there are always large spaces reserved for the development of new connections. Moreover, points which are already involved in a definite conditioned activity frequently change their physiological rôle and become connected with some other activity of the animal.
The idea of the cerebral cortex as a mosaic of functions is in part Indicated in the current anatomical and physiological teaching. The structural complexity of the specific receptor organs of special Sense, the rich complexity of the cellular structures of the cortex, and the complexity of their distribution conform quite readily to the idea of the mosaic character of the central nervous system. The rather rough localization of function which has been achieved During the last century of experimental physiology is no less in accord with such a conception. The final resolution of the many delicate problems involved in this conception is, however, a complex and difficult matter; up to the present it has been possible only to make the very first attempts towards an [p. 220] experimental study of the simplest aspects of the problem, and even these aspects demand the utmost skill and resource of inquirer.
The existence of a localization of function in the cortex is in some cases Sufficiently proved by the evidence of structure alone, definite receptor elements having been shown to stand in connection with definite cortical cells. The study of conditioned reflexes has shown, however, that a conditioned reflex appears at first in a generalized form, i.e. that excitation irradiates from its point of initiation to embrace also cells belonging to other receptor elements beyond the boundary of the area of the cortex primarily connected with the stimulated receptor; the problem becomes still more complicated when we take into account also the subsequent concentration of excitation upon its point of initiation.
Two different problems present themselves for consideration. In the first place the question arises how the functional demarcation of two positive cortical points each connected with a different unconditioned reflex is effected, and in the second place the similar question arises as to the mechanism of the functional demarcation of: neighbouring positive and negative cortical points belonging to the unconditioned reflex. The study of anatomical localization of function is obviously of no assistance in solving these problems. The investigation of the first question is being conducted at the present time, and therefore cannot yet be discussed; our investigations up to the present have been confined almost exclusively to the second problem, since it is the simpler.
We shall commence with a simple and obvious case. A positive conditioned alimentary reflex was established to a tactile stimulation of the skin on the right shoulder, and a negative conditioned reflex to a similar stimulation of the skin on the right thigh. After these reflexes had been thoroughly established, the effect of a stimulation of other places on the skin was tried. The different sites of stimulation were: (1) the front paw 17 cms. below the positive point on the shoulder; (2) the side of the animal 12 cms. caudally from the positive place on the shoulder; (3) the side of the animal 15 cms. in front of the negative place on the thigh; and (4) the hind paw 18 cms. below the negative place on the thigh. The salivary secretion was measured during 30 seconds of isolated stimulation of each of the above places. The results are summarized in the following table : [p.221]
Similar results were obtained in two other dogs [experiments of Dr. Foursikov].
It can be seen that under the influence of two definite external stimuli, which affected the animal under opposite conditions (one reinforced by the unconditioned reflex and the other remaining without reinforcement), two perfectly definite and separate points within the cutaneous analyser were given, the one excitatory, and the other inhibitory, properties. Around each of these points there was established a corresponding region of positive or negative influence, these regions of positive and negative influence extending towards each other, although each maintaining its distinctive properties. The smallness of the positive reflex on stimulating the place on the side of the animal nearer to the thigh, and the absence of positive reflex on stimulating the hind paw, indicate the predominance of inhibitory properties in these respective points. In the absence of a nucleus of inhibition the positive effect due to the initial generalization would have spread over the whole of the analyser in such a manner that the decrement of the positive reflex with increase of distance from its point of initial development would have been gradual, as was shown in the tenth lecture.
The experiments described in this lecture show that external stimuli evoking antagonistic processes provide the first method by which a functional mosaic character of the cortex originates. Many other examples of this can be found in the lecture upon the analysing function of the cortex.
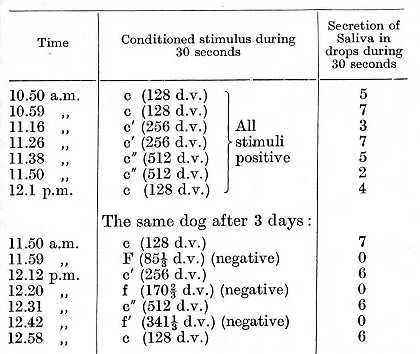
More complicated experiments were conducted upon three more dogs. In two dogs different tones [experiments of Dr. Siriatsky], and in the third dog tactile stimulation of different areas of the skin [experiments of Dr. Koupalov], were used as conditioned stimuli, [p. 222] alternate areas being respectively positive and negative. The object of the experiments was to determine the mode of development of the mosaic character, the stability and the delicacy of its pattern, the interaction of its different points, the effect of stimulation of the spaces intermediate between the positive and negative places, and the effect of development of such a functional mosaic upon the general condition of the animal. Some of the results of these experiments will be given now, others will be described in further lectures, although it should be mentioned that the investigations are still being pursued. In one dog all the tones C of five neighbouring octaves of an organ (64-1024 d.v.) were used as positive conditioned alimentary stimuli, while all the tones F (851/3-13651/3 d.v.) were inhibitory conditioned stimuli. In the following tables the respective C's and F's are designated according to Helmholtz's scale. The following experiment illustrates the final result obtained with one dog:

In the second dog different tones from f (170 2/3 d.v.) to f’ sharp (360 d.v.) were used in alternate order as positive and negative conditioned stimuli. e' sharp (335 d.v.) was given the properties of an excitatory stimulus, e' (320 d.v.) properties of an inhibitory stimulus, c' (256 d.v.) excitatory, a (213 1/3 d.v.) inhibitory, g (192 d.v. excitatory and f (170 2/3 d.v.) inhibitory, these tones being enumerated in descending order. As can be seen, the intervals between the respective positive and negative tones were in this dog irregular, and smaller than in the first dog. The final stage of development of the reflex is shown in the following experiment: [p. 223]

In the third dog the conditioned alimentary reflexes were tactile. The apparatuses used for the stimulation were always arranged in strictly identical positions along a line extending along the left side from the hind leg along the whole length of the body to the left leg, the apparatuses being set at a distance of 12 cms. from each other measured from the centre of one apparatus to the next. The stimulated places are numbered in order from No. 1, the left hind leg to No. 9, the left fore leg. Stimulations of the places represented by odd numbers were given excitatory properties, and stimulations the places represented by even numbers inhibitory properties. The following is a typical experiment:
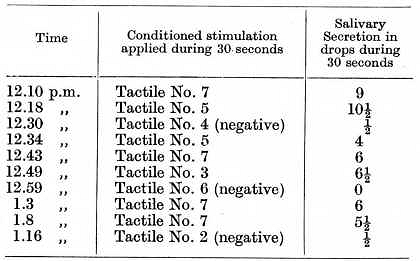
The development of this functional mosaic of the cortex presented at the beginning some difficulty, but with continuation of the experiments it grew progressively easier. The reflexes were developed not simultaneously but in succession. One of the most interesting [p. 224] points brought out by these experiments was the spontaneous development in the first and third dogs, in the final stages, of some new reflexes, and it is important to note that the new conditioned properties in these cases developed within the region of the antagonistic nervous processes. In the case of the first dog the positive conditioned reflex to c" was of very long standing. In one experiment this tone c" evoked a secretion of 8 drops during 30 seconds, while on the contrary f" even on its first application failed to produce any positive effect. This inclined us to regard f" as inhibitory from the start, and the truth of such an assumption was definitely proved in an experiment on the following day, when, f" being used as the first stimulus, it not only remained without any positive effect but also left a considerable inhibitory after-effect, so that the positive reflex to the succeeding stimulus c" became very much diminished. It is obvious therefore that f" assumed its inhibitory properties spontaneously, without requiring any contrasting, but to all appearance simply in virtue of its situation in the neighbourhood of the positive cortical point corresponding to c". The same phenomenon repeated itself with c"', which when tested for the very first time gave a positive effect apparently in virtue of its proximity to the negative f".
In the third dog, with tactile conditioned reflexes, the places Nos. 1 and 9 also developed their full positive effect spontaneously without any previous reinforcement, but most probably in virtue of being within the sphere of influence of the previously established negative places Nos. 2 and 8.
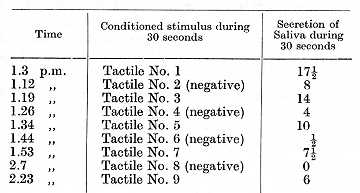
Clearly these facts should be interpreted as due to mutual induction initiated by the pre-existing points of excitation and inhibition in regular alternation. These experiments show us why any rhythmic activity is performed more easily and less exhaustingly than an arhythmic one. The experiments on all three dogs showed us that in the case of a regular alternation of positive and inhibitory stimuli all the reflexes were exceedingly precise. In some cases such a regular alternation leads even within one experimental day to a more and more precise functional localization of positive and negative cortical points. This can, of course, most clearly be demonstrated in cases where at the beginning of the experiment the effect of different points was lacking in precision. The following results were obtained with the dog in which tactile stimuli were employed as the alternating excitatory and inhibitory conditioned stimuli; the [p. 225] experiment was performed before the negative stimuli became well established:
The following is another example of an experiment on the first dog taken at the time when the reflexes were not yet quite regular:
The above experiments indicate that the mutual induction of the antagonistic nervous processes of excitation and inhibition should be regarded as the second contributing factor to the development of a mosaic of functions in the cerebral cortex. [p. 226]
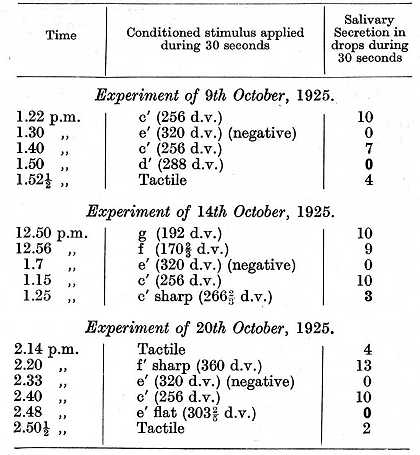
The same dogs were used to determine the extensity of the excitatory and inhibitory processes around the respective points of stimulation, and to determine also whether there existed any mosaic of neutral territory interwoven with the mosaic of excitation and inhibition. For this purpose, in the dogs in which a mosaic of tones was established, other tones were used intermediate between the tones which had been given positive or negative significance, and the effect produced by these intermediate tones was observed. A positive effect, in the form of a salivary secretion, was a direct evidence of the tone having definite excitatory properties. An absence of secretory effect, on the contrary, gave no special indication whether the tone had any inhibitory conditioned properties or was merely neutral, and special experiments had to he made to test whether such tones left any inhibitory after-effect or exhibited any effect of positive induction. In the following experiment, between the positive tone 256 d.v. and the negative tone 320 d.v. three tones were taken: 266 2/3, 288 and 303 2/5 d.v. respectively.
 [p. 227]
[p. 227]
The above experiments show that the tones intermediate between positive c' and the negative e', namely, the tone d' and the semitones c' sharp and e' flat, assumed a different significance. The semitone c' sharp produced a definite though comparatively small positive effect, i.e. it still belonged to the excitatory region of c' ; on the other hand the tone d' and the semitone e' flat at the first glance were equal in their effect, since both gave a zero secretion. It was only by testing their inhibitory after-effect that a difference between these two tones could be revealed. The conditioned tactile stimulus when applied two minutes after the use of the semitone e' flat diminished in its effect by 50%, i.e. it was under the influence of the inhibitory after-effect. When, however, tested under precisely the same conditions after the tone d' the tactile stimulus gave a full positive effect. The semitone e' flat, therefore, belonged unquestionably to the region of the inhibitory e' ; the tone d', however, either was entirely neutral or else carried much weaker inhibitory properties, an alternative which can be finally settled only by more delicate experiments. The whole question of the possible existence of completely neutral points in the cortex is being further investigated.
It was mentioned in the first lecture and again in the beginning of the present lecture that additions can always be made in any pattern of the functional mosaic both with respect to its extent and its complexity. Moreover it is subject also to frequent reconstruction, one and the same point changing its physiological significance and becoming connected successively with different physiological activities of the organism. In respect to this we have only one series of experiments, conducted by Dr. Friedman, as follows:
One and the same agent served at first as a conditioned alimentary stimulus, later it was transformed into a conditioned stimulus for acid; reversely, old conditioned stimuli to acid were transformed into alimentary ones. Two dogs were used for experiments with the first set of transformations, and a third dog was used for the second set. The transformation of conditioned stimuli was effected by the substitution of one unconditioned reflex for the other. The following is the general sequence of events: The conditioned stimulus, on transition from one unconditioned reflex to another very quickly, even within a single experimental day, loses its secretory effect and gives a series of zeros for a comparatively long time; only after [p. 228] considerable practice with the new unconditioned stimulus does it gradually resume excitatory properties, and these now belong to a conditioned reflex based on the new unconditioned one, as shown by the very definite change in the composition of the saliva secreted by the submaxillary gland and in the character of the motor reaction. The complete replacement of the one conditioned reflex by the other required about 30 reinforcements by the new unconditioned stimulus. After a considerable practice of the conditioned reflexes to acid the conditioned stimuli were once more transformed back again into alimentary ones; the transformation occurred rapidly and only a few reinforcements were needed. indicates that the original alimentary connection was still preserved in spite of the establishment of a new connection with the reflex to acid.
However, the main interest of the experiments of Dr. Friedman lay in the investigation whether a differentiation which had been established for the conditioned stimulus to one reflex would be preserved after its transformation. A detailed description of an experiment will be given in respect to one dog only, since the experiments with all three dogs gave identical results. The alimentary conditioned stimulus in this dog was given by the tone of 2600 d.v. ; a precise differentiation from this tone was established to the neighbouring tone of 2324 d.v. The positive conditioned alimentary reflex was now transformed into a conditioned reflex to acid, and when the differentiation was tested it was found from its very first application to be complete and to exert the same inhibitory effect as before. A new and more precise differentiation was now established on the basis of the "acid" reflex to a note a semitone higher than the original positive tone, i.e. 2760 d.v., and it was found that when the tone of 2600 d.v. was transformed back again into a conditioned alimentary stimulus this new and finer differentiation also retained its precision.
In the course of the preceding lectures we had occasion to refer to the fact that one and the same point of the cortex could, in accordance with the given experimental conditions, be a point of origin either of excitation or of inhibition. These transformations of the physiological significance of a definite cortical point are attained with comparative ease and are effected with considerable rapidity both in respect of positive and of negative stimuli (experiments with induction, p. 194), and in respect of their ultimate con- [p. 229] nection with one or another unconditioned reflex (experiments just described). However, amongst our material we have a number of cases in which the definite positive or negative quality of some region of the cortex, or even a temporary correlation between the two processes, assumed an exceedingly persistent character which could be changed only with great difficulty, or even failed to be changed. These cases were met with in the experiments of Dr. Frolov upon conditioned positive and negative trace reflexes. The results of these experiments were of such unique character that it is necessary to describe them in detail. In the case of one dog a trace conditioned reflex to acid was established to a tone of 1740 d.v. produced by an organ pipe: the tone was continued for 15 seconds and the acid was administered after a pause of 30 seconds. This trace reflex was repeated 994 times in the course of one year and nine months. Its latent period, counting from the beginning of the tone, was equal to very nearly 25 seconds (10 seconds reckoned from the cessation the tone). After the termination of the investigation originally planned it was resolved to abolish the trace character of the reflex and to give it the character of a delayed reflex, evoking the positive secretory effect during the action of the tone, and not some time its termination. For this purpose acid was administered at 15th second after the beginning of the tone. After twenty reinforcements with this new interval it was still found that, not only did the reflex fail to appear during the 15 seconds of the isolated action of the tone, but even the administration of acid at the 15th second evoked a secretion only after the old latent period of 25 seconds from the beginning of the tone. The use of other auditory stimuli in place of the tone did not alter the result, and when the tone and the other auditory stimuli were protracted for 45 seconds without the administration of acid the secretion always started at about the 25th second from the beginning and then increased rapidly, giving about 10 drops during the remaining 20 seconds of isolated action of the conditioned stimulus. After this failure to diminish the latent period, the stimulus was reinforced practically simultaneously, i.e. two seconds after the beginning of the tone preceding the reinforcement. Nevertheless, even after several score of repetitions the conditioned secretion started only after a latent period of 20-25 seconds from the beginning of the tone ; this latent period could not be changed whether the acid was administered at the 2nd second or whether the tone was continued singly for a much greater [p. 230] length of time. The last experiment performed in this series is given in the following table:

A conditioned alimentary reflex to a tone f sharp of a tone variator was now developed in this dog, but the latent period of this reflex also could not be reduced below 24 seconds, and it was only when a tactile stimulation was used as a conditioned stimulus and reinforced with acid after 2 seconds that a closer approximation to a simultaneous alimentary reflex was at last obtained. At the twenty-fourth test the reflex was measured by 12 drops during 30 seconds, with a latent period of 2 seconds.
A similar persistence of a thoroughly established cortical inhibition was observed in the case of another dog. The trace of a tone of 1740 double vibrations produced by an organ pipe was used as a conditioned inhibitor to the positive conditioned stimulus of a metronome beating at a rate of 104 per minute. The trace of the tone even after one minute still exerted a full inhibition upon the reflex to the metronome. Such an inhibition had to be developed of course gradually, starting with recent traces of only a few seconds. The experiments were conducted with this conditioned inhibition for well over two years, and the tone, besides serving as a conditioned trace inhibitor, was also differentiated from other tones, which, of course, led to an extreme intensification of its inhibitory properties. At the termination of these experiments the animal was used for another research in which it was desired to develop a positive conditioned reflex to a sound of a microphone which happened to be practically identical in pitch with the tone which had served as the conditioned inhibitor in the first series of experiments. The positive conditioned reflex refused for a long time to develop, and when the [p. 231] metronome was tested shortly after the sound of the microphone had been applied a most powerful conditioned inhibition was revealed. The experiments were now modified so that the inhibitory combination of the microphone with the metronome was directly reinforced by the unconditioned stimulus; in other words, a procedure was adopted which exactly reversed that employed for the development of the conditioned inhibition to the original trace of the tone. The inhibitory combination now quickly acquired excitatory properties, and somewhat later the sound of the microphone applied singly acquired a positive significance of its own. However, the tone of the organ pipe which had been originally developed as the conditioned inhibitor did not acquire a positive character; neither did it lose its inhibitory properties : these had to be destroyed by the same method as was used for the destruction of those of the microphone, but the process occupied a much longer time.
These cases of extreme stability of the inhibitory process must be regarded as exceptional. It is very probable that the intensity of inhibition in these cases was favoured by some special conditions of the experiments, and it is possible that this exceptional persistence of the inhibitory process can be correlated with the experiments upon hypnotism and sleep which will be described further on.
Along with these cases of an extremely stable " inhibitability " of definite points of the cerebral cortex must be placed a case of a similarly stable excitability. In an experiment by Dr. Bierman a well-established conditioned alimentary reflex, to the tone of 256 double vibrations produced by an organ pipe, was differentiated from twenty-two separate and distinct tones ranging in pitch from 768 d.v. to 85 d.v., the tone of 258 d.v. (c') being always accompanied by food, while all the others remained unreinforced. The positive tone was contrasted in each experiment several times with one or other of the negative tones. In this manner the positive cortical point became surrounded by a series of inhibitory points. However, this apparent encircling of the positive point by inhibitory ones did not lead to a diminution of the effect of the positive tone ; on the contrary it led to its extreme intensification. When in subsequent experiments the animal was experimentally subjected to a profound physiological sleep, so that, for example, shrill whistling and banging on the door of the experimental room containing the dog did not awaken it, the positive conditioned tone awakened the animal immediately and [p. 232] evoked a full conditioned secretion. Experimental proof will be given in further lectures that sleep itself is nothing but a form of internal inhibition, and from this point of view the experiment just described must be regarded as a case of extreme stability of the excitatory process in a definite point of the cerebral cortex which successfully resists the surrounding inhibition.
The foregoing discussion of the experimental evidence permits us to regard the activities of the cerebral hemispheres as a true mosaic of functions. All the numerous individual cortical points, each at any definite moment, have some very definite physiological significance, while the whole mosaic of functions is integrated into a complex dynamic system and perpetually achieves a unification of the individual activities. Every new localized influence playing upon it influences to a greater or less extent the entire system. Consider, for example, a dog which possesses at any given moment a definite number of conditioned reflexes. The addition of new positive, and especially of new negative, reflexes exercises, in the great majority of cases an immediate, though temporary, influence upon the older reflexes [experiments of Dr. Anrep]. Further, even when no new reflexes are added, and it is only a rigidly adopted sequence of their order which is changed, their magnitude undergoes distinct diminution, showing a considerable predominance of inhibition. In the following experiment by Dr. Soloveichik a dog was subjected at intervals of 10 minutes to the conditioned alimentary stimuli of a metronome, an electric lamp, a whistle and a tactile stimulus, always repeated in this order in every experiment. The following table gives the results of the final experiment with this order of the stimuli :
 [p. 233]
[p. 233]
In an experiment on the following day the order of the stimuli was changed.

The diminution in the magnitude of the conditioned reflexes observed when the order of their administration is changed sometimes appears immediately, as in the foregoing experiment, and in other cases becomes more obvious in the succeeding experiment when the stimuli are again applied in their usual order. The diminution continues for several days, and then the reflexes quickly return to their normal magnitude. Such a diminution I the strength of the reflexes as occurred towards the end of the first experiment given above (3.59, 4.9, 4.19 p.m.) is of frequent occurrence and will be more fully discussed in the next lecture.