
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3173
(Return to index)
By Ivan P. Pavlov (1927)
Translated by G. V. Anrep (1927)
Posted December 2000
The development of inhibition in the cortex under the influence of conditioned stimuli.
Conditioned stimuli, acting as they undoubtedly do through the intermediation of definite cortical cells, provide the obvious means whereby the physiological characteristics of these cortical cells can be studied. One of the most important of these properties is that under the influence of conditioned stimuli they pass, sooner or later, into inhibition. In the previous lectures upon internal inhibition it was shown that in all cases when a positive conditioned stimulus repeatedly remains unreinforced, it acquires inhibitory properties, i.e. the corresponding cortical cells enter under its influence into a state of inhibition. The present lecture will be devoted to the study of the intimate mechanism of this phenomenon, and of the part played therein by the unconditioned reflex and by other conditions which retard or accelerate the development of this inhibitory state.
The transition of the cortical cells into an inhibitory state is of much more general significance than could be inferred from the facts which have been discussed up to the present, concerning the development of internal inhibition. The development of inhibition in the case of conditioned reflexes which remain without reinforcement must be considered only as a special instance of a more general case, since a state of inhibition can develop also when the conditioned reflexes are reinforced. The cortical cells under the influence of the conditioned stimulus always tend to pass, though sometimes very slowly, into a state of inhibition. The function performed by the unconditioned reflex after the conditioned reflex has become established is merely to retard the development of inhibition.
The following is the most commonly occurring example of this phenomenon: We are dealing, we will suppose, with a conditioned reflex which is delayed by 30 seconds, i.e. a reflex in which the [p. 235] conditioned stimulus acts singly for exactly 30 seconds before the addition of the unconditioned stimulus. Let us suppose that when the reflex is well established its so-called latent period -- the interval time from the beginning of the conditioned stimulus to the onset secretion -- is equal to five seconds. Now this latent period remains practically unaltered for a certain length of time, which varies greatly in different dogs. As time goes on the latent period lengthens out, and finally during the 30 seconds of the isolated action of the conditioned stimulus no trace of salivary secretion is produced. It is, however, only necessary to delay the administration of the unconditioned stimulus by a further 5-10 seconds for secretion again to be obtained during the prolongation of the isolated action of the conditioned stimulus. On continuing the experiment for some time as before, i.e. with a delay of 30 seconds, and then again introducing a delay increased by a further 5-10 seconds, no conditioned secretion is obtained any more. In order to obtain a conditioned secretion the administration of the unconditioned stimulus must be delayed for a still greater length of time. Finally a stage is reached when no conditioned secretion can be obtained during any length of isolated action of the conditioned stimulus. This gradual disappearance of the conditioned secretion in reflexes with a constant delay occupies very different periods of time in different dogs. In some it takes only days or weeks, and in others it takes several years. The conditioned secretion disappears later with tactile, than with thermal, later still with visual, stimuli and latest of all with auditory stimuli, especially if the latter are discontinuous.
The following is an example of the relative differences between the various conditioned stimuli with respect to the rapidity of transition of the cortical elements into an inhibitory state under the influence of a definite delay [experiments of Dr. Shishlo]. The first conditioned stimulus which was developed in a particular dog was one belonging to the tactile analyser: the administration of food was usually delayed 10 seconds from the beginning of the tactile stimulus, but on rare occasions a 30 seconds' delay was introduced. The reflex first appeared at the 27th stimulation, and within five weeks, after 179 stimulations, the reflex became stable at 8 drops during one minute. The development of a second alimentary conditioned reflex to a thermal cutaneous stimulus of 45° was now commenced, the normal period of delay being as before 10 seconds. This reflex [p. 236] developed quickly, and when at the twelfth stimulation a delay of 30 seconds was introduced a secretion of 4 drops was obtained. On continuation of the usual reinforcement applied 10 seconds after the beginning of the thermal stimulus the reflex rapidly diminished in strength and at the 33rd test, when the delay was specially prolonged to one minute, only one drop of secretion was obtained.
The following observation leaves no doubt that the disappearance of the conditioned reflex, notwithstanding its invariable reinforcement, is an expression of a progressive development of inhibition in the cortical elements. When an effective positive conditioned stimulus is applied shortly after the application of a conditioned stimulus which has, as described, just lost its positive properties, the resulting reflex suffers a diminution. Similarly, when one among a number of conditioned stimuli has lost its positive effect its disuse in the experiments leads to an increase in the effect of the remaining stimuli. To my mind such results can only be interpreted on the assumption that real inhibitory properties have been acquired by those stimuli which have lost their positive effect. This phenomenon should not be confused with what was previously described as the result of internal inhibition of delay. The inhibition of delay is revealed by the so-called latent period which is observed in every conditioned reflex and which remains unchanged for a considerable period of time. The phenomenon which is being described here, on the other hand, is characterized by its invariable progressiveness. The inhibitory state of the cortical elements under the influence of conditioned stimuli develops more quickly with longer delayed reflexes; the longer the isolated application of the conditioned stimulus the quicker the development of the inhibition. For example, it often happens that a reflex with a delay of 10 seconds remains unchanged in its strength for a very considerable time and thus permits of exact experimentation; the same reflex when delayed to 30 seconds quickly becomes unsuitable for experiments through the progressive development of inhibition. Such a case is taken from experiments by Dr. Petrova:
A dog has a conditioned alimentary reflex established to the sound of a metronome, and throughout the whole period of work the reflex which has been delayed for 10 seconds remains constant in strength. The following experiment has been chosen at random : [p. 237]
As soon as the isolated application of the conditioned stimulus is prolonged to 30 seconds the reflex becomes inconstant in strength, and on repetition diminishes to zero during the time of one single experiment.

The above two experiments were conducted with an interval day.
In view of the great variations existing between individual dogs it has been found very useful, and sometimes essential, to employ in different dogs, reflexes which are delayed by different lengths of time. It now becomes obvious why the long-delayed reflexes develop only with difficulty and why during the beginning of our work they could be obtained only in some of the dogs. Moreover, it becomes easy to understand why in many of the experiments previously described the positive alimentary conditioned reflexes diminish during a single experiment and in some cases after only a single application -- as in the experiment on page 232 of the previous lecture. This diminution is due to the repetition of the conditioned stimulus and not to any other factor, such, for example, as a gradual satiation of the animal during the experiment in the case of alimentary reflexes. The latter is obviously not the case, since on repeating any one of the established conditioned stimuli, only the effect pro- [p. 238] duced by that one becomes diminished, while other conditioned stimuli may preserve their full effect up to the very end of the experiment. When a definite positive conditioned stimulus has already shown a tendency to assume inhibitory properties, then after a short interval in the experiments, or even at the beginning of a day's experiment, its first application produces a considerable secretion, but on repetition the stimulus quickly diminishes in its effect and becomes inhibitory in spite of its being reinforced at every application.
It is in the interest of the experimenter for most of the experiments to have at his disposal reflexes of a constant intensity. To obtain such reflexes it is necessary in many cases to fight against the progressive tendency of conditioned reflexes to undergo inhibition. At first on purely empirical grounds, and subsequently more rationally, a number of ways of combating this inconvenience were evolved. For obvious reasons the more effective of these methods were such as established conditions exactly the reverse of those which led to the progressive diminution of the conditioned reflexes. Foremost among such methods was the introduction of an occasional abrupt shortening of the length of isolated action of the conditioned stimulus. If, for example, the reflex originally has been delayed 30 seconds, the practice is now adopted of reinforcing at the 3rd to 5th second. Of course, during this short period of time the conditioned stimulus cannot evoke a measurable reflex, and very often no salivary secretion whatever can be observed during so short a period of delay. The short delay, however, is only introduced as a temporary expedient, being, so to speak, only a therapeutic measure applied for the purpose of regenerating the conditioned reflex. Afterwards, when the usual 30 seconds' delay is restored, it is found that the reflex is as strong and as constant as in the beginning. It is useful to make this return from the short-delayed reflex to the long-delayed one by stages, only gradually increasing the length of the isolated action of the conditioned stimulus. The efficacy of this method, measured by the permanence of the reconstituted reflex, depends upon the of weakening of the conditioned reflex, upon the time during which this weakness is allowed to persist, and upon the length of time during which the short delay is practised as a therapeutic measure. A short period of practice restores the reflexes only to a small degree and for a short time. Where a profound weakening of the reflex has been allowed to persist for a very long time, the method which has [p. 239] just been described, and which is, generally speaking, very effective, no longer suffices, as will be shown presently, to restore the reflex.
A conditioned reflex which has become weakened in the course of long practice can be helped towards recovery by avoiding any numerous repetitions of the conditioned stimulus within a single experiment, and, if possible, confining the use of the stimulus to single applications. A similar beneficial effect results from a simple interruption, even for a few days, in the work upon the weakened conditioned reflexes.
Besides these there are several subsidiary methods, as, for example, to increase the strength of some of the conditioned stimuli or to increase their number, by the summation of positive induction, or finally by increasing the strength of the unconditioned stimuli. At present, however, interest centres mainly in the direct methods referred to above.
There are cases where the extent of the diminution in the strength of conditioned reflexes is such that none of the above methods can be of any help -- all positive conditioned reflexes simply disappear. The animal grows inert in the stand during the experiment, and even declines the food which is given after application of the conditioned stimulus. This can be observed even with dogs which have served, on account of the stability of the conditioned reflexes, for extremely exact experimentation during a period of many years.
What is to be done with an animal in such a condition? In the earlier period of our work such a dog would undoubtedly have been discarded as one which could be of no further use for our purpose. Now, on the other hand, the condition of such a dog is regarded as calling for further investigation, which, as a matter of fact, can easily be carried out. It is sufficient to stop the use of all the old conditioned stimuli, and to develop instead conditioned reflexes to new stimuli, for the seemingly insurmountable difficulty to disappear. The new conditioned reflexes develop extremely quickly, and this is not surprising since all the extraneous reflexes which originally interfered with the development of conditioned reflexes have long since disappeared. The newly developed reflexes quickly attain a maximal constant and strength, and the animal entirely returns to its original condition and can be used for further experimentation. In view of extreme importance of this fact I shall describe in greater detail the history of one of the dogs [experiments by Dr. Podkopaev].
Experiments with this dog were started in June, 1921. One after [p. 240] another different positive and negative conditioned reflexes were established. Among these were several reflexes to tactile stimulation of different places of the skin (designated by numbers in the succeeding tables). All the reflexes were extremely constant in their magnitude and in the length of their latent period, and the dog was used during several years for experiments on various problems. The following example illustrates the strength of the positive reflexes as tested in an experiment of 30th August, 1922:
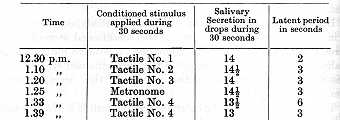
An experiment performed on the 6th August, 1923, involving various positive stimuli and a conditioned inhibitor gave the following results:

An experiment of 12th June, 1924, with the positive reflexes and a negative one (conditioned inhibition, but with a different inhibitor) resulted as follows:
 [p. 241]
[p. 241]
Towards the end of 1924 the stimuli very often had a zero effect and the dog no longer took immediately the food which was presented. Several expedients were tried in order to get rid of this increasing inhibition. Instead of tactile stimuli auditory ones were mainly used, and instead of long delays very short ones; intervals of as much as a month and a half were made between experiments; an increase in the strength of the unconditioned stimuli was tried; and the animal was kept on the floor during the experiments, instead of in the stand. Any beneficial effect produced by these methods was only fleeting: the animal became more and more languid and often altogether declined food given after the application of conditioned stimuli. This State of the animal persisted throughout the whole of the year 1925. At the end of 1925 the use of all the old conditioned stimuli was abandoned and the new ones were introduced. This procedure led to a quick and definite change in condition of the animal. It again became alert, and immediately took food on presentation at the end of the application of the new conditioned stimuli. The conditioned reflexes were quickly established and reached a constant and considerable strength, while the latent periods returned to normal. These results remained steady in subsequent experimentation. The following is an early experiment during this phase, performed on the 21st January, 1926. The reflexes in this experiment are delayed by 15 seconds, and an attempt is made to reinstate among the new conditioned stimuli one of the old ones to the sound of a metronome.

The differences in the magnitude of the conditioned reflexes in the above experiment depend to a certain extent on the different stages of development which they have reached.
The general significance of these experiments is obvious. The isolated action of the conditioned stimulus, even though followed by [p. 242] the unconditioned, leads to the development of a state of inhibition in the cortical elements, and this development is the quicker greater the length of isolated action of each single conditioned stimulus and the more often such a stimulus is used. It thus becomes apparent that the difference between the process of development of inhibition as studied in the early lectures with respect to different cases of internal inhibition and in those cases which have just described is not of fundamental importance; although in most cases very substantial it is obviously only one of degree. In those cases in which the conditioned stimulus remains unreinforced the inhibitory process develops very quickly; in those cases with reinforcement which have just been described the development of the inhibitory process is usually delayed -- sometimes so considerably that its development may even remain unsuspected. Only in very rare cases is the rate of development of inhibition in both groups of experiments nearly or completely identical.
We now come to the problem as to the mechanism by which unconditioned stimuli retard the development of the type of inhibition which we are now considering. In the second lecture we saw conditioned reflexes do not develop when the unconditioned stimulus precedes the neutral agent which is required to be made into a conditioned stimulus. This is probably a result of external inhibition, the strong excitation produced by the unconditioned stimulus leading to inhibition of that cortical area which is but weakly excited as a result of the application of the neutral stimulus. If this be the case, the question arises whether the unconditioned stimulus similarly produces external inhibition of the cortical areas corresponding to an already established conditioned reflex. To test this point experiments were performed as follows:
A firmly established conditioned stimulus was superimposed on the background of the reinforcing unconditioned stimulus, i.e. the unconditioned stimulus was applied first and the previously established conditioned stimulus was applied only after the effect of the unconditioned stimulus had become apparent. This mode of experimentation was continued for several weeks or months on many dogs. The results were without exception uniform: the conditioned reflex never kept its original strength, but either weakened considerably or else disappeared altogether.
The stronger and the more practised the conditioned reflexes were, the more slowly they disappeared under this treatment, [p. 243] whereas weak and recently established reflexes lost their conditioned properties very quickly. The following experiment by Dr. Solovaichik gives an illustration of this case.
The dog employed had many strongly developed alimentary conditioned reflexes, each of which gave upwards of 20 drops of saliva during 30 seconds.
The establishment of a new reflex to a hissing sound was now begun. This reflex developed very quickly, and counting from the sixth application to the eleventh it gave secretions of 10, 8, 13, 9, 9 and 10 1/2 drops during 30 seconds. Altogether only eleven reinforcements were made, after which the conditioned stimulus of the hissing sound was superimposed upon the unconditioned. The application was repeated in this way fifty-four times in the course of thirty-two successive days. The following table shows the trial of the hissing sound after the fifty fourth superimposition:
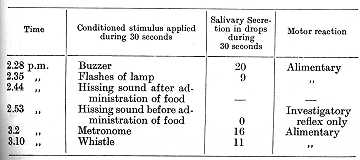
It is thus clear that the administration of the unconditioned stimulus results in an inhibitory state of those cortical elements on which the conditioned reflex depends --these cortical cells becoming temporarily unresponsive to their normal exciting stimulus. The above experiment does not, however, disclose the inner mechanism determining the difference in rate of development of inhibition of the cortical elements in the case of reinforcement and non-reinforcement.
The sum of all the periods of action of a positive stimulus required for the development of inhibition without reinforcement is usually incomparably less than that of the periods of isolated action of the same stimulus with reinforcement. It follows therefore that the transition of the excited cortical elements into an inhibitory state [p. 244] is not determined by the aggregate duration of the isolated stimulus. It is, however, probable that the excitation in the case of non-reinforcement persists for a long time after the cessation of the isolated action of the conditioned stimulus, while in the case of reinforcement the excitation is curtailed from the beginning of the unconditioned stimulus. The aggregate duration of the excitation produced by the stimulus may, therefore, be the determining factor, but other explanations are possible and the matter cannot be decided without further experimentation.
The fundamental fact in all these experiments, which repeats itself time after time, is the transition sooner or later into inhibition of the state of the cortical elements acted upon by the conditioned stimulus. So far as concerns all the experimental evidence at our disposal up to the present, this transition must be regarded as depending on a functional exhaustion of the cortical elements as a result of their activity in response to a stimulus. Such an exhaustion would obviously be dependent upon the duration and intensity of this activity. On the other hand it is also obvious that the process of inhibition cannot be regarded as identical with such functional auto-destruction of the cortical elements, since a state of inhibition which is initiated in an active cell spreads to other cortical elements which were not active and which were not therefore functionally exhausted.
The rate at which cortical elements become subjected to inhibition in the case without reinforcement fits in with the extreme sensitivity which they exemplify in their extreme need for constant nutrition, being, as is well known, finally and irreparably destroyed by an arrest of the blood supply far sooner than any other tissue of the body. It is in complete harmony also with the conception of the cerebral cortex as a signalling apparatus. The fact that the unconditioned stimulus which is signalled induces during its action an inhibition in the cortex is only an artistic finishing touch to the efficiency of the machine. I may permit myself to use the analogy of an efficient and watchful signalman who after having performed his responsible duties has to be provided with an immediate rest during which he is refreshed, so that he may afterwards perform his task again with the same efficiency as before.
Another question arises whether there exists such a minimal period of isolated action of a conditioned stimulus as does not lead in time to a progressive development of inhibition in the cortical [p. 245] elements. The details of this problem are receiving a full experimental investigation in our laboratories at present, but there are some experiments already performed which have a probable bearing upon it. In dogs which had had for a very considerable time conditioned reflexes delayed by 30 seconds, and in which the conditioned stimuli still retained their full effect, all the reflexes were transformed into almost simultaneous ones by reinforcement 1 to 2 seconds after the beginning of the conditioned stimulus. This modification in the procedure immediately began to reflect itself on the secretory and motor components of these reflexes, bringing about also a disturbance of the previous balance in strength between positive and negative conditioned reflexes. The magnitude of the positive reflexes increased considerably, while the negative reflexes were to a large extent dis-inhibited; in other words excitation began to predominate over inhibition [experiments of Dr. Petrova and Dr. Kreps]. The following is the method used by Dr. Kreps:
The dog, in view of an especially exaggerated tendency to inhibition, was not kept in its stand during the experiments, but was kept on the floor. Among other conditioned reflexes this dog possessed a positive alimentary reflex to 132 beats per minute of a metronome, while a rate of 144 beats per minute served as the stimulus to a precise and stable differentiation. A considerable time before the experiment a conditioned inhibition had been established in which flashes of a lamp acted as the conditioned inhibitor to a rate of 120 beats per minute of the metronome. The conditioned inhibition was absolute, but had not been used for a considerable time. All the reflexes were delayed 30 seconds. These delayed conditioned alimentary reflexes were now transformed into simultaneous ones, the food being presented one second only after the administration of the conditioned stimulus. The modification in the mode of reinforcement led to a disappearance of the conditioned inhibition, which could not be re-established in spite of 100 applications of the inhibitory combination during 36 days. The differentiation of the metronome, which was absolute before the transformation of the reflexes into simultaneous ones, was also dis-inhibited, and in the succeeding 33 repetitions which were performed within 13 days the differentiation continued to be unsatisfactory (3 drops as against 6 to 7 of the positive reflex). On returning to the delay of 30 seconds with all the reflexes, the differentiation again became absolute during the very first day, and the conditioned [p. 246] inhibition was completely re-established after 3 days, as shown by the following table:

On transforming the reflexes again into simultaneous ones the conditioned inhibition was almost completely dis-inhibited after only three repetitions of the simultaneous reinforcement.
It remains still to be determined whether in the experiments just described, in which the conditioned stimulus is almost immediately followed by the unconditioned, there will be no tendency towards a progressive development of inhibition with prolonged practice, or whether the apparent vigour of the reflexes will be only temporary and will, though much later, nevertheless be superseded by inhibition.
Hand in hand with the exhaustion of the cortical elements there goes of course their recovery. We should expect, therefore, that the inhibition which appears to stand in some kind of relation to functional exhaustion of the cortical elements should disappear with their functional recovery. This expectation fits the case of spontaneous recovery of extinguished conditioned reflexes which after some interval of time return to their normal strength. Regarded from this point of view it becomes more easy to understand how the slowly developing inhibition on repetition of the reinforced conditioned reflexes is replaced by a temporary return of the delayed reflexes to their original strength when a period of very short delays is introduced or when a considerable interval is made between experiments. By shortening the period of isolated action of the conditioned stimulus in the one case, or by completely avoiding for a time any repetition of the excitation in the other case, the functional exhaustion of the cortical elements is diminished and a better opportunity is afforded for complete recovery.
Experiments are now being performed with the object of specially [p. 247]
studying the restoration of conditioned reflexes in the case of experimental extinction, and in the case of the gradual spontaneous development of inhibition occurring in spite of reinforcement. An example of such an experiment may be taken from a research by Dr. Speransky.
Amongst others the following serve as positive conditioned alimentary stimuli: beats of a metronome, intensification of the general illumination of the room, the sound of a whistle, and the appearance of a circle. The acoustic reflexes were somewhat stronger than the visual ones, giving 10 to 12 drops during 30 seconds preceding reinforcement while the visual ones gave only 6 to 8 drops. The stimuli were all applied at intervals of 10 minutes and always in the above order. In every experiment all the stimuli were repeated once or twice. Following a series of experiments of this type the next experiment would consist in stimulation by the metronome alone. Twelve successive applications were made at intervals of 10 minutes, each being reinforced. The first two applications gave 12 and 11 drops of salivary secretion respectively during their isolated action, while the last two gave 9 drops each. It is thus seen that simple repetition of the stimulus led to a diminution in its effect by 25%. On the following day a similar experiment was conducted, but the interval between the stimuli was made very much shorter, being only 1 1/2 minutes. The use of such short intervals was possible only because in this particular dog the secretion caused by the reinforcement with food finished extremely quickly and, as had been found by previous determinations, well within the 1 1/2 minutes. In this variation of the experiment the reflex which measured 11 drops on the first stimulation became diminished to 4 1/2 drops on the third stimulation (diminution by 60%), to 2 drops on the 8th stimulation (diminution by 82%); passing with more or less regular undulations of 2, 5, and 7 drops, it gave on the 22nd stimulation only one drop and then no secretion at all, the dog even refusing food after the last three stimuli. When the visual stimulus of increased illumination of the room was applied 1 1/2 minutes after the last stimulation by the metronome a reflex of 2 1/2 drops of secretion was produced, and the animal took the food. Moreover, when food was given without a previous conditioned stimulus the animal devoured it with avidity. On the next day a return was made to the original method of application of all the conditioned stimuli at intervals of 10 minutes. The metronome was the first stimulus to be applied and gave 6 1/2 drops. [p. 248]
On introducing a second stimulation by the metronome as the 5th stimulus the secretion obtained was only 1 1/2 drops. Finally, introduced a third time as the 9th stimulus, the metronome did not produce any secretion at all. In this experiment the secretion given by the other stimuli, although slightly below normal on their first application, did not greatly diminish on repetition. The reflex to the metronome was found to be restored to its usual intensity on the following day, and now it showed no further tendency to diminish on repetition. The foregoing experiment presents many points of interest. It is seen that one and the same stimulus which is constantly reinforced and repeated many times at long intervals of time loses only little of its effect: the same stimulus when applied at short intervals of time quickly diminishes in its effect at first, then after a wave-like variation it ends by giving a complete zero of secretory and motor reactions, the animal declining food after the conditioned stimuli. In spite of this, another and usually much weaker stimulus immediately evokes both secretory and motor effects. The fact that the animal in the stand consumed with avidity a large amount of food when not preceded by the conditioned stimulus shows that satiety of the animal played no part. Relating the effect of the beats of the metronome to different states of the cortical elements it must be concluded that in the case of frequent stimulation these elements get functionally exhausted and have insufficient time for recovery, so that after a preliminary oscillation which may be regarded as a struggle between excitation and inhibition the cortical elements pass completely into inhibition. The weaker visual stimulus diminishes considerably in its effect after the disappearance of the conditioned reflex to the stronger stimulus of the metronome. On the following day the cortical elements which are stimulated by the metronome are still not fully recovered and pass again into inhibition when the stimulus is repeated. This inhibition is of the same character as the internal inhibition which has been described in previous lectures, and it exhibits the same properties of irradiating to other cortical elements which were not primarily involved. The experiment shows that the inhibition irradiating to neighbouring cortical elements standing in connection with other conditioned stimuli leads to a diminution of their positive effect, exactly as was described for internal inhibition. That the inhibition in the neighbouring cortical elements is really due to an irradiation of the primary inhibition is evidenced by the fact that on repetition of the stimuli they do not show such a [p. 249] rapid diminution in their effect as is observed in the case of the metronome. On the third day the cortical elements related to the metronome became almost entirely functionally restored.
The observations described in this lecture open up many important problems, and in the first instance the question of rapidity of recovery from the inhibitory effect of different stimuli during complete rest and during activity, the effect upon this recovery of reinforcement, so on. Besides these comparatively straightforward problems there are more complex and more difficult ones. It is obvious that only certain cases of the development or disappearance of inhibition can be brought into relation with a supposed functional exhaustion and recovery of the cortical elements, and we cannot interpret in this fashion the cases of permanent and unvarying inhibitions in which the activity of the cortex is so rich -- for example, all cases where an established inhibitory conditioned stimulus evokes an inhibition of the cortical elements directly and without a preceding phase of excitation -- as, for instance, in the case of differentiation conditioned inhibition. The problem presented by inhibitions of the latter type becomes still more complicated when we remember that those points of the cortex which become the centres of such direct inhibitions are never transformed into centres for excitation even though the experiments are interrupted for weeks and months.