
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3713
(Return to index)
CONDITIONED REFLEXES: AN INVESTIGATION OF THE
PHYSIOLOGICAL ACTIVITY OF THE CEREBRAL CORTEX
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE XV
Internal inhibition and sleep as one and the same process with regard to their intimate mechanism.
In the last lecture we arrived at the very important conclusion that under the influence of our conditioned stimuli the cortical elements invariably enter sooner or later into an inhibitory state. With frequent repetitions of the stimuli this happens extremely quickly, and it may legitimately be regarded as an expression of the fact that the cortical elements, which represent the highest point of development of the nervous system, are extremely sensitive and therefore are functionally exhausted with comparative ease. The progressively developing inhibition, which itself cannot be regarded as a functional exhaustion, but which is a result of exhaustion, assumes the rôle of a protector of the cortical elements, preventing any excessive fatigue or dangerous functional destruction of this highly sensitive structure. During the period when the cells are in a state of inhibition, being free from activity the cortical elements recover their normal state. This applies to all the cellular structures of the cortex equally, and therefore under conditions in which a great number of cortical points are repeatedly entering into a state of excitation the whole of the cortex may be expected sooner or later to become subjected to inhibition. Such a state of widely spread inhibition actually does occur, exactly in the same manner as in the case of individual cortical elements, and is familiar to all of us as the common and everyday occurrence of sleep. The complete and continuous proof of this contention is spread over the whole of our twenty-five years' work upon the hemispheres, and at the present time no part of the physiology of the hemispheres studied by the method of conditioned reflexes is better substantiated. Drowsiness and sleep were met with in our experimental animals from the very beginning of our work, and we have been obliged to direct our attention towards them continually. This, of course, has led to the collection of an immense number of facts, which were bound in different phases of our experimentation to receive many [p. 251] different interpretations. However, already for many years all these varied interpretations have been fused into a final one harmonizing with all the facts at our disposal. This conclusion is, in essence, that sleep and what we call internal inhibition are one and the same process.
The fundamental condition of the appearance and development of internal inhibition and sleep is exactly the same. It consists in the more or less prolonged or many times repeated isolated action of a conditioned stimulus producing stimulation of the cellular structures in the cortex. In all cases of internal inhibition which were discussed in the fourth to the seventh lectures drowsiness and sleep were met with continually. In the case of extinction of a conditioned reflex some animals even at the first extinction showed not only a disappearance of the conditioned secretory and corresponding motor reaction but also a great dullness as compared with the normal state of the animal before the extinction. Repetition of extinctions, in the course of a number of days, even if all the conditioned stimuli were reinforced in between, led in every case to an obvious drowsiness and even sleep of the animal in its stand, though no such symptoms had ever previously been observed. The same happens, but to a much greater extent, in the development of differentiation. To take an example. An animal has conditioned reflexes established to different stimuli, including one to a definite musical tone. During the whole period of work the animal remains alert. The development of a differentiation of a tone close to the positive one is now started, and it is noticed that during the process the animal gets drowsy. The drowsiness gradually increases, and often culminates in a deep sleep with a complete relaxation of the skeletal muscles, and snoring, so that when now other positive conditioned stimuli are administered and reinforced by food it is necessary to stir up the animal and even to introduce the food forcibly into its mouth to initiate the act of eating. Exactly the same thing happens in the case of development of long-delayed reflexes (for example, with a delay of three minutes), and in the early period of our work this interfered with our researches, for, being not yet thoroughly familiar with the technique, it was impossible to obtain in some animals the reflex we required, on account of the development of sleep. The same happens also in the development of conditioned inhibition, but to a smaller extent.
In all the foregoing cases of internal inhibition sleep develops [p. 252] fairly rapidly, which depends on non-reinforcement of the conditioned stimulus. In the case of the slowly developing internal inhibitions produced by the repeated use of reinforced conditioned stimuli over a period of months or years, the development of sleep is proportionately slower and generally speaking stops short at one or other of the intermediate stages between the alert state and sleep itself, depending on the dog employed. In this respect the animals differ exactly as they differ in respect of rapidity of development of the common forms of internal inhibition.
It is not necessary to give any examples of individual experiments upon this transition of internal inhibition into sleep, since all our experiments abound with observations showing that internal inhibition invariably passes into sleep unless special precautions are taken. As there is practically no stimulus of whatever strength that cannot, under certain conditions, become subjected to internal inhibition, so also there is none which cannot produce sleep. Very powerful electric shocks applied to the skin, when used as conditioned alimentary stimuli, led, after many months of use in the experiments of Dr. Eroféeva, to a progressively increasing internal inhibition in spite of continuous reinforcement, and in the experiments of Dr. Petrova they became most effectual agents in inducing sleep. Similarly, different external agencies in their role of conditioned stimuli fall into an identical order of classification as regards the rapidity with which they lead to internal inhibition and to sleep. It was mentioned in the preceding lecture that internal inhibition develops most readily with thermal and least readily with auditory stimuli; in exactly the same manner sleep develops quickly with thermal conditioned reflexes and more slowly and less frequently with auditory conditioned reflexes. The interference by sleep in the case of thermal conditioned stimuli was indeed so persistent and upset the work to so great an extent that in the early period of our research I had real difficulty in finding collaborators who would agree to work with these stimuli.
Finally, the length of isolated action of the conditioned stimulus was mentioned as a factor determining the development of internal inhibition; so also it is a factor in determining the development of sleep. In some dogs, while the conditioned reflex was delayed only 10 or 15 seconds, the animal remained fully alert during experiments spread over years, but so soon as the reflex was delayed for 30 seconds drowsiness and sleep appeared. The results of this type of experiment [p. 253] are often truly striking, for the quick transition from full alertness into true physiological sleep, due to this seemingly insignificant change in the experimental conditions, is amazing. Examples with variable length of delay and with variable precision of results are strewn over our work in abundance.
All those methods described in the preceding lecture as retarding or abolishing that progressive growth of internal inhibition, which develops in the case of frequently repeated conditioned reflexes notwithstanding their constant reinforcement, can be used with equal efficiency for the purpose of resisting sleep.
At this point the following question naturally arises: If sleep closely coincides in its appearance and disappearance with internal inhibition, how is it that the latter plays such an extremely important part during the alert state of the animal, serving for the most delicate physiological mechanism of equilibration of the higher organism with its environment? To my mind all the facts which have been given in the preceding lectures dispose at once of the apparent contradiction. Internal inhibition during the alert state is nothing but a scattered sleep, sleep of separate groups of cellular structures; and sleep itself is nothing but internal inhibition which is widely irradiated, extending over the whole mass of the hemispheres and involving the lower centres of the brain as well. Thus internal inhibition in the alert state of the animal represents a regional distribution of sleep which is kept within bounds by the antagonistic nervous process of excitation. Such a restricting antagonism has been illustrated already in the lectures upon the functional mosaic the cortex and upon its analysing activity.
In the case of extinction the development of sleep is prevented only if after extinction the conditioned stimuli are systematically reinforced and extinction is not repeated too often. In differentiation of stimuli the developing internal inhibition--which tends at first be accompanied by sleep -- can only be definitely restricted within its own analyser by inserting the inhibitory conditioned stimuli between repeated applications of the positive conditioned stimulus. In this manner the process of excitation which is repeatedly being evoked antagonizes a universal spreading of internal inhibition. Exactly the same is observed in the case of conditioned inhibition and inhibition of delay. In all these cases if the experiment is conducted with forethought drowsiness and sleep appear only as phasic events during the time when the physiological demarcation [p. 254] between the areas of excitation and inhibition is not yet fully established. However, as soon as the conditions of the experiments lead to a prevalence of inhibition sleep again reappears. The following is a striking example: In the lecture upon the functional mosaic a dog was mentioned in which a tone served as the stimulus to a positive conditioned reflex, while twenty neighbouring tones up and down the scale were differentiated as stimuli for negative conditioned reflexes. This animal was never inclined to drowsiness or sleep when a balance was maintained between the number of repetitions of the positive and negative conditioned stimuli, and under these conditions the dog always gave full reflexes to the applications of the positive stimulus. As soon, however, as the inhibitory tones were used several times in succession, the dog quickly fell into such profound sleep that even most powerful extraneous stimuli failed to awaken it. When a return was made to the interposition of the negative conditioned stimuli between applications of the positive one, sleep was never observed to develop. In this respect, such experiments as have already been mentioned in connection with the mosaic character of cortical functions -- especially those with the use of tactile stimuli -- are very instructive. In spite of a tendency of tactile conditioned stimuli to favour the development of drowsiness and sleep, the dog used by Dr. Koupalov never exhibited any signs of drowsiness, although a conditioned tactile mosaic was practised in this dog for over two years (p. 223). This was obviously due to the inhibition being constantly limited and checked in its irradiation by the antagonistic excitatory process. Another method of combating any wide irradiation of inhibition is to increase the number of positive conditioned stimuli, and so check the spread of inhibition from its initial points of development.
The following observation of Dr. Petrova, which to some extent also bears on this subject, is more complicated. A long-delayed alimentary reflex to a metronome was being developed without the previous establishment of a simultaneous reflex: the length of the delay was 3 minutes. During several days the dog became more and more drowsy, and finally fell into a state of profound sleep. Obviously the inhibition which developed during the first period of action of the metronome was so powerful that it prevented the development of the phase of excitation which normally precedes reinforcement. Five new agencies were now used to develop further alimentary conditioned reflexes -- these agencies together with the original [p. 255] stimulus were allowed to act for only 5 seconds before reinforcement. Drowsiness quickly disappeared and all the reflexes developed with ease. The delay before reinforcement was now gradually extended in all six reflexes by 5 seconds each day. Corresponding with the increase in delay before reinforcement the latent period of the reflexes became longer, and finally without any interference of sleep six long- delayed reflexes became firmly established, all with a preliminary period of inhibition of 1-1/2 minutes. The process of excitation, originating in six different points of the cortex, allowed the inhibition to develop only gradually, limiting it both in time and space and preventing the development of sleep.
To the same group as the last belong some further observations which were made in only a few dogs, all of them very easily subjected to inhibition. These dogs developed drowsiness and sleep simply on account of limitation of movements when placed in the stand. Sleep could be avoided, at least for some length of time, by conducting the experiments with the animals free upon the floor. It is probable that under the latter condition stimuli originating within the motor apparatus and the skin provided fairly regular foci of excitation within the cortex which counteracted to some extent any wide irradiation of conditioned inhibition. However, another factor of probably greater importance also undoubtedly played a part, and to this we shall return later.
What has been shown to take place in the cortical elements with respect to the development of internal inhibition under the influence of conditioned stimuli, can be observed to the same extent in the case of stimulation of the cortical elements by agents which have no special conditioned physiological significance. As has previously been mentioned, among the different reflexes the investigatory reflex has a special importance. This reflex has a point d’appui in the cells of the cortex as well as in the lower parts of the brain. In the normal animal the reflex is undoubtedly produced with the active co-operation of the cortex. This view is supported by the exquisite sensitivity of the reflex, for it is evoked by any minutest change in the environment. This is made possible only through the presence of the higher analysing activity of the cerebral cortex and is wholly unattainable by the lower parts of the brain alone. The investigatory reflex as we know, invariably weakens on repetition, and finally disappears altogether. Special experiments conducted in my laboratory by Professor Popov showed that the disappearance [p. 256] of the investigatory reflex is based on the development of inhibition, and is in all details analogous to extinction of conditioned reflexes.
If the agent which is responsible for the investigatory reflex ceases, on repetition at frequent intervals of time during one single experiment, to call forth the corresponding motor reaction, a prolongation of the interval in the same experiment restores the reaction exactly as in the case of extinguished conditioned reflexes. Similarly, a definite investigatory reflex which has only just disappeared on account of repetition of the stimulus becomes temporarily re-established by an application of some new extra stimulus calling forth another investigatory reflex. It follows, therefore, that the extinguished investigatory reflex undergoes dis-inhibition exactly as do the underlying positive reflexes in cases of internal inhibition. If the investigatory reflex to a definite agent is repeatedly evoked in the course of a number of days it permanently disappears, just as does a systematically non-reinforced conditioned reflex. Finally, such an extinguished investigatory reflex can be temporarily re-established by administration of stimulants (caffeine) exactly, for example, as in the case of conditioned differentiated reflexes (p. 127). The inhibition of the investigatory reflex invariably leads to drowsiness and sleep (even more easily than the inhibition of conditioned reflexes). In the following experiments by Dr. Chechoulin the development of inhibition and sleep in the case of the investigatory reflex was studied by means of conditioned stimuli.
The dog used for these experiments had a conditioned alimentary reflex established to a whistle. Hissing, bubbling, tactile stimulation of the skin and other stimuli were all applied for the first time to bring about the investigatory reflex.
Now, beginning from 4.21 p.m., the bubbling sound was repeatedly applied during periods of 30 seconds and at intervals of 2 minutes. During the first three applications there were movements of orientation of the animal in the direction of the sound, and these movements [p. 257] gradually became weaker. With the fourth application the first signs of drowsiness made their appearance. Up to the eighth repetition the sleep was interrupted at different moments of the stimulation. During the eighth and ninth stimulations all movements of the animal disappeared. At 4.43 p.m. the sound of bubbling was applied for 10 seconds, and then the whistle was added and kept on for 30 seconds. This brought about neither motor nor secretory reaction and the sleep continued. Administration of food awakened the animal; it took the food but even after that it still remained drowsy. The experiment with the conditioned stimulus continued as follows:
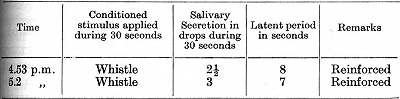
It should be remembered that this dog was never observed to fall asleep in its stand during the usual experiments with conditioned reflexes. In the succeeding experiments new agents bringing about an investigatory reflex were repeated up to the point of sleep, or sometimes only until the stage of disappearance of the motor reaction. Twenty-one days after the experiment recorded above tactile stimulation of the skin was used as the extra stimulus for the investigatory reflex, the experiment proceeding as follows:
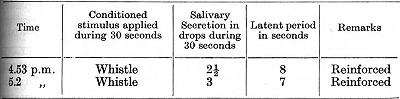
The conditioned reflex in the above experiment was twice reinforced after only five seconds of the action of the stimulus in order to maintain the normal strength of the reflex to the end of the experiment.
Starting from 4.25 p.m. an application of the tactile cutaneous stimulus was made during 30 seconds and similar applications were repeated at intervals of one minute. During the first three applications the animal turned its head towards the place on the skin where [p. 258] the tactile stimulus was applied. During the fourth and fifth repetitions there were no movements at all, but there was no drowsiness of the animal. Now, at 4.32½ p.m., the tactile stimulus was applied singly during 10 seconds, after which the whistle was added to it and both stimuli were continued together during 30 seconds. Fifteen seconds after the beginning of the action of the whistle there was a commencement of salivary secretion, but during the whole remaining period of stimulation only 2 drops were recorded. The experiment continued as follows:

The experiments show that on repetition the motor component of the investigatory reflex gradually diminished, and that, on con- continuing the repetitions further, drowsiness developed and became more and more profound, although in some experiments before drowsiness appeared the stimulus remained, during a certain interval of time, apparently without effect. Nevertheless, superimposition of a conditioned stimulus upon the extraneous stimulus showed that during the whole period of this apparent ineffectiveness the investigatory agent was exerting an inhibitory influence (the experiment with the tactile stimulus as extra stimulus). This inhibition of the conditioned stimulus was not due to external inhibition, for, so far from producing inhibition of the conditioned reflexes, a weakened investigatory reflex actually dis-inhibits them (compare the sixth lecture for the action of the investigatory reflex upon the two phases of delayed conditioned reflexes). It is obvious therefore that inhibition and sleep develop as a result of the repetition of the investigatory reflex, in the former case leading to a diminution and in the latter to a disappearance of the conditioned reflex (the experiment with the bubbling sound).
The same is very well shown also when the experiments are conducted on puppies [experiments of Dr. Rosenthal]. On monotonous repetition of a stimulus under constant environmental conditions the puppies fall asleep very quickly and often with surprising simultaneity. It is, I think, a common experience that man, when unused to an intensive mental life, usually falls into drowsiness [p. 259] and sleep when subjected to the accompanying monotonous stimuli, however unfortunate such drowsiness or sleep may be as to place and as to time. This means, of course, that the definite cortical elements which react to such protracted external stimuli become functionally fatigued and pass into a state of inhibition, which in the absence of a counteraction by an excitation of other places, spreads over the hemispheres and leads to sleep. The extreme rapidity with which the cellular structures of the cortex undergo functional fatigue and become subjected to inhibition can be contrasted with the persistence of function of the cellular structures of the spinal cord and medulla under identical conditions. Experiments by Dr. Zeliony in our laboratory showed that while in a normal dog an investigatory reflex to a definite sound quickly vanished, the same sound in a dog with extirpated cortex, under identical conditions, called forth an investigatory reflex in a stereotyped manner and for an unlimited number of times.
To return again to conditioned reflexes. The development of inhibition with its ultimate expression in the form of sleep is due to functional fatigue of the cellular structures of the cortex. This is borne out by the following observations which were made repeatedly in our laboratory. In dogs in which any analyser had been surgically damaged, positive conditioned stimuli related to this analyser could scarcely be continued even for a very short time singly, since they tended quickly to assume inhibitory properties; quite often they never excited any preliminary positive action at all, but behaved as inhibitory stimuli from the start. This phenomenon is especially constant and easy of demonstration in the case of the damaged cutaneous analyser. After extirpation of the gyri coronarius and ecto-sylviusanterior (see Fig. 8), positive reflexes to tactile stimulation of the extremities, pelvis and shoulder became replaced for several months by inhibitory reflexes. That the reflexes were now truly inhibitory was proved by the fact that positive conditioned reflexes related to other analysers showed their full normal effect before the application of a tactile stimulus, but diminished in their positive effect or lost it altogether after the application of the tactile conditioned stimulus. At the same time tactile stimuli quickly and easily produced sleep even in dogs which never before the operation fell asleep in response to tactile stimuli. These facts often assumed the following extremely impressive form. A tactile conditioned stimulus which was applied to the part of the cutaneous surface related [p. 260] to the damaged portion of the analyser would lead to inhibition and sleep, but the same stimulus applied to the skin corresponding to the non-damaged portion of the analyser would give a full positive effect, leaving the animal fully alert [experiments of Dr. Krasnogorsky, Dr. Rosenkov and Dr. Archangelsky]. To the same group of observations belong those made in our laboratory during the period of shortage in Russia a few years ago. The semi-starved animals could not be used for experiments with conditioned reflexes, since all positive conditioned stimuli assumed inhibitory properties, and the dogs invariably developed sleep exactly in conjunction with the application of the conditioned stimuli. Obviously the general malnutrition of the dog had powerfully affected the functional resistance of the cortical elements [experiments of Dr. Frolov, Dr. Rosenthal and others].
In the examples enumerated up to the present we have had only cases of transition of inhibition into sleep, but the reverse can also take place, sleep passing into inhibition. To take an example. A conditioned reflex delayed by 3 minutes has been established. The animal is placed in the stand and is fully alert, but so soon as the conditioned stimulus is applied the animal becomes drowsy and no salivary secretion is evoked during the whole 3 minutes. When food is given at the end of the third minute the animal takes it but slowly and reluctantly. The stimulus is repeated several times in the same experiment with the usual variations of the interval between the applications; at each stimulation the dog becomes more alert and the secretion appears at first towards the end of the 3rd minute. On further repetition of the stimulation the secretion augments and finally the three-minute period of stimulation divides itself approximately into two equal parts. In the first part there is no secretion, although the animal remains entirely alert ; in the second part there is a copious secretion, and at the end of the stimulation the animal takes the food promptly and eats it with avidity. In this case the widely irradiated inhibition (sleep), which appeared in the beginning on account of the preponderance of the inhibition initiated during the first part of the action of the conditioned stimulus, gets gradually concentrated into a restricted inhibition. This concentrating of the inhibition is brought about through the influence of the progressively increasing excitation determined by the second part of the action of the conditioned stimulus and by the dis-inhibitory effect of reinforcement. [p. 261]
It sometimes happens that the reverse, namely a pure replacement of inhibition by sleep, is obtained with the long delay of 3 minutes, or even with delays so short as 30 seconds. The animal, which has previously kept fully alert in its stand during the experiment, now falls asleep, each time exactly at the beginning of the action of the conditioned stimulus. The eyes close, the head droops, the whole body relaxes and hangs on the loops of the stand, and the animal emits an occasional snore. After the lapse of a definite period of time -- in the short delay 25 seconds or in the case of the long delay 1 1/2-2 minutes -- the animal quickly and spontaneously awakens and exhibits a sharp alimentary motor and salivary reaction. It is clear that in this case an inhibition which is generally concentrated becomes replaced by diffused inhibition, i.e. sleep.
Finally, it can also be shown [experiments of Dr. Foursikov] that a summation of two distinct and different inhibitions leads to sleep. For example, the dog has a well-established long-delayed conditioned reflex to a metronome: the length of the delay is 3 minutes. No salivary secretion occurs during the first two minutes of the stimulation, but at the end of the second minute the secretion appears, and reaches a maximum towards the end of the third minute. An extraneous stimulus of a weak hissing sound is now made to accompany the conditioned stimulus. The hissing sound dis-inhibits the inhibitory phase of the reflex, while a small motor reaction in form of an investigatory reflex towards the hissing is observed. The conditioned reflex is reinforced. On a repetition of this combination not only does the investigatory reflex to the hissing disappear, but the alimentary reflex also, and the animal becomes obviously drowsy. This experiment can only be interpreted in the following way: The investigatory reflex to hissing undergoes extinction on its first application, and now, therefore, the hissing sound initiates an inhibitory process. This inhibitory process summates with the inhibitory phase of the delayed reflex and strengthens it to such an extent that the excitatory phase of the reflex is never allowed to develop, being replaced by general drowsiness of the animal. That this is the true interpretation of the experiment appears abundantly in the sequel. On the next repetition of the conditioned stimulus, and without the addition of the weak hissing sound, a regular delayed reflex, with its two phases well pronounced, is obtained, and on repeating after this the combination of the metronome with the hissing sound the conditioned reflex again [p. 262] disappears and obvious drowsiness takes its place. The following are the actual figures of the experiment:
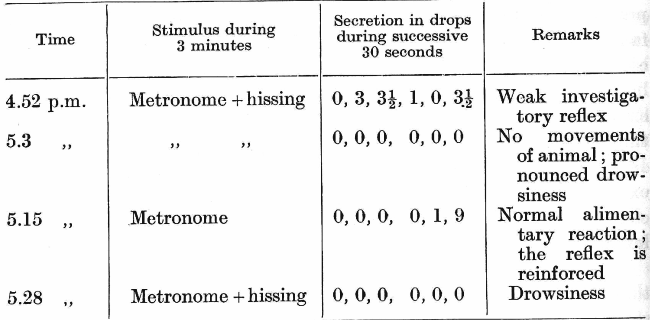
In this connection I conceive it useful to draw attention to the following interesting point. It is evident that the above experiment, together with that of Dr. Chechoulin mentioned previously (p. 256), reveals still another phase in the action of extra stimuli upon conditioned reflexes. A powerful extra stimulus, as will be remembered from the sixth lecture, at first brings about through the investigatory reflex a complete inhibition of the delayed reflex. On repetition, when the investigatory reflex considerably weakens, it brings about only dis-inhibition of the first phase of the delayed reflex. Finally, as we have just learned, the extra stimulus yet again inhibits the reflex, but now by another mechanism: it becomes itself a stimulus for a direct initiation of an inhibition in the cortex. A weak extra stimulus, as was just shown by the experiment of Dr. Poursikov, brings about at first a weak and transitory investigatory reflex and so leads on its very first application to dis-inhibition of the delayed reflex (4.52 p.m.). Afterwards, the weak extra stimulus itself initiates a second, and now direct, inhibition.
The following general properties of inhibition and sleep also uphold our view as to their identity. In the preceding lectures abundant evidence was given to establish the fact of irradiation and concentration of the inhibitory process within the mass of the cerebral cortex, and it was shown that the development of the inhibition was extremely slow, being measured by minutes. Moreover, it varied in respect to rate in different animals and under different [p. 263] conditions. There is no question but that sleep also does not develop instantly. We know from our own experience how drowsiness and sleep overtake us only gradually, and how sometimes they spread only slowly and with difficulty; and some investigators, indeed, have endeavoured to study experimentally the problem of the gradual involving of the activity of the different sense organs, and its more complex mental concomitants. We know also how variable is the rate of transition in human beings between sleep and waking, and the same variability has been observed in our experimental animals. Moreover, during our lectures it has constantly been mentioned how inhibition, which at first develops with difficulty, gets reproduced with greater and greater ease upon practice and repetition and by using different forms of inhibition. Exactly in the same manner, extra stimuli, and conditioned stimuli which upon repetition bring about a state of sleep, with practice bring about this state more and more easily.
The following is of special interest. As was discussed previously, inhibition induces excitation. Corresponding with this, in some animals in which the inhibitory phase of a delayed reflex is replaced by sleep, this appearance of sleep is on some occasions preceded by a short period of a slight but definite general excitation of the animal. The phenomenon is still more obvious and constant when sleep is induced under the action of a repeated and prolonged neutral extra stimulus. It was often observed in the experiments by Dr. Rosenthal, that, when the neutral stimulus evoked definite drowsiness in a puppy, and before the animal completely fell asleep, it passed through a fleeting phase of excitation, moved about uneasily, scratched itself, and barked without any obvious reason, holding its nose up into the air. A similar state of general excitation preceding sleep, as is well known, often occurs in children. It is legitimate to regard such phenomena as an effect of induction. The excitation in initial stages of anaesthesia could also perhaps be interpreted from this point of view.
I believe the aggregate of facts given in the present lecture can be taken as sufficient proof of the view that sleep and internal inhibition are fundamentally one and the same process. I personally do not know, up to the present, of a single fact in all our researches which contradicts this conception. It is to be deplored, however, that we have as yet no reliable graphic method of registration of sleep. On some occasions we tried to apply for this purpose a [p. 264] graphic registration of the position of the head of the animal. A perfection of some such method for the graphic registration of sleep is greatly to be desired, so that the whole evidence regarding sleep can be expressed in an exact quantitative manner. The details of our normal everyday existence are in full agreement with the foregoing interpretation of sleep. Our daily work, for some of us a round of exceeding monotony and for others extremely rich and varied, in either case must in the end determine an appearance of sleep. A prolonged stimulation of one and the same point in the cortex leads to a great and profound inhibition, and this irradiates widely so as to involve the whole of the cortex and the lower parts of the brain. In the case of a varied activity, although no given point of the cortex attains such a profound depth of inhibition, yet the great number of inhibitory points leads to a widely distributed inhibitory state even without wide irradiation, and this also descends to affect some of the lower centres of the brain. Of course a great number of quickly changing stimuli following in succession may often exert a very prolonged and powerful resistance to the general dissemination of inhibition over the hemispheres, thus delaying the onset of sleep. A well-established rhythm in the changes from wakefulness to sleep and from sleep to wakefulness may determine a beginning of sleep even without a sufficient functional fatigue of the cellular structures of the cortex. Both cases have had sufficient illustration in our experiments, in the analogous relations between the excitatory and inhibitory processes.