(Return to index)
Posted March 2001
[Classics Editor's note: Pavlov used both square and round brackets
in his texts. These have been preserved but can lead to confusions as to
which insertions are the author's and which are the editor's. Page numbers,
reference numbers, and the occasional "sic" have been inserted in
square brackets by the Classics editor. All other insertions (e.g., on
p. 31) are by Pavlov. -cdg-]
LECTURE XVIII
Pathological disturbances of
the cortex, result of functional interference (continued).
In the present lecture we shall discuss further experiments and observations upon the pathological states of the cerebral cortex. The inquiry is one of particular interest, not only on account of the special attention devoted to this subject at the present time, but also on account of several fortuitous occurrences. It has become possible to trace how, as a result of different injurious influences, the activity of the cortex gradually and by scarcely noticeable stages deviates from normal and becomes pathological; often also the pathological states can be made use of for inquiry into nervous processes taking place under purely physiological conditions, since under pathological conditions different aspects of the nervous processes which are screened off from us by the unified and balanced complexity of the normal physiological state become dissociated or accentuated. In the lecture upon the hypnotic states occurring under normal conditions it was mentioned that the most interesting of these were subjected to experimental investigation only after they had previously been observed in exaggerated form in a pathological case.
In one of our dogs which, it will be remembered (p. 270), had several positive and negative conditioned reflexes, a tactile stimulation at the rate of 24 per minute had been established as a positive stimulus and a stimulation at the rate of 12 per minute as a negative one: moreover, as usually under normal conditions, the relative intensity of the positive reflexes was directly determined by the relative strength of their conditioned stimuli. In one of the experiments the positive rate of the tactile stimulation was made to follow the inhibitory rate without any interval of time. This apparently small modification of the experiment was sufficient to create a pathological disturbance of the cortical activities in this dog. At first, for a few days, all positive conditioned reflexes disappeared completely; this was followed by a series of different modifications [p. 302] in the strength of the reflexes, a state of affairs which lasted for many days, the relation between the magnitudes of the reflexes and the intensities of the stimuli gradually changing from one phase to another. The entire disturbance lasted for 5 1/2 weeks, after which the reflexes returned to normal. Obviously this dog falls into one group with the inhibitable dog (described at the end of the preceding lecture), in which functional interferences with the cortical activities produced a profound nervous disturbance which was accompanied by a disappearance of all positive conditioned reflexes for many months. In the dog just described the same type of disturbance was produced, but lasted for only thirty-six days, within which period the activity of the cortex passing through different stages finally returned to normal.
It becomes clear on considering all the pathological cases so far described, that the underlying cause of their development is in every instance the same. Broadly we can regard these disturbances as due to a conflict between the processes of excitation and inhibition which the cortex finds difficult to resolve.
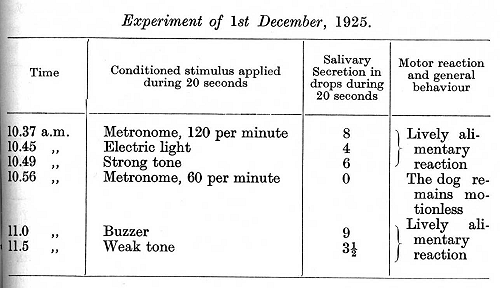
Besides these pathological cases we have at our disposal others which present features of perhaps even greater interest, firstly on account of certain intrinsic peculiarities, secondly on account of peculiar features in the mode of their origination. The case which is to be described first was studied from day to day during many months, and in view of the considerable interest it presents the description will be given in detail [experiments of Dr. Rickman]. The experiments were carried out on a dog which was exceedingly inhibitable. The animal had served in the laboratory for different experiments for a very long time, and it had among others an inhibitory alimentary conditioned reflex to a metronome rate of 60 beats per minute, while a rate of 120 beats per minute served as a positive stimulus. The strength of the various positive reflexes was definitely related to the strength of their respective conditioned stimuli. The inhibitory conditioned reflex, which at the time of the experiments had been repeated 266 times, was constant and precise, being so concentrated that its inhibitory after-effect upon the positive reflexes was very short. The strength of the reflexes during this normal period is represented in the table shown on the opposite page.
The dog belonged to a group which can be called "expert at inhibition," in view of the fact that in this group all the types of [p. 303] internal inhibition develop with great ease and precision. We now determined to investigate in further experiments the degree of stability of the inhibitory process in this dog. For this purpose it was decided to transform the negative conditioned stimulus into a positive one by the method which is usually the most effective, namely, repeated reinforcement of the inhibitory stimulus without any intermediate introduction of the positive stimuli (see p. 198). However, the destruction of the inhibition in this dog proceeded extremely slowly. The hitherto inhibitory stimulus was followed by reinforcement 4-7 times in succession on each of three successive
days. The first sign of any destruction of the inhibition was observed at the seventeenth application with reinforcement, in the form of a very small secretion without any accompanying alimentary motor reaction. At the twenty-seventh reinforcement the salivary secretion was already considerable. No definite disturbances in the other positive stimuli could be observed at this period, excepting a certain tendency to equalization of the secretory effect of strong and weak positive conditioned stimuli. The experiment of the 14th December on page 304 shows the strength of the reflexes at this time.
The secretory reaction thus established to the rate of 60 beats of the metronome did not, however, remain constant, but in spite of continued reinforcement quickly declined and at its thirtieth repetition fell to zero. Moreover, it was now noticed that immediately after an application of the metronome at the rate of 60 practically none of the positive reflexes could be elicited (18th December). [p. 304]


In its general behaviour the animal appeared to be perfectly healthy and when free on the floor it consumed the same food as that offered during the experiment with great avidity, just as it did in the above experiment after the conditioned stimulus applied before the metronome (12.4 p.m.).
In succeeding experiments, though some of the positive effect of the metronome rate of 6O per minute returned, its strong inhibitory effect upon other conditioned reflexes nevertheless continued as before.
In all those experiments, however, in which the metronome was not used all the reflexes were perfectly normal, excepting that the weaker stimuli gave a somewhat smaller secretion towards the end of an experiment than usual.
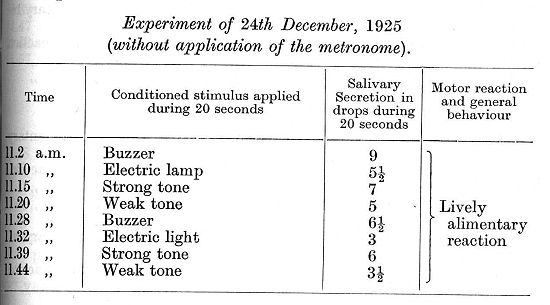
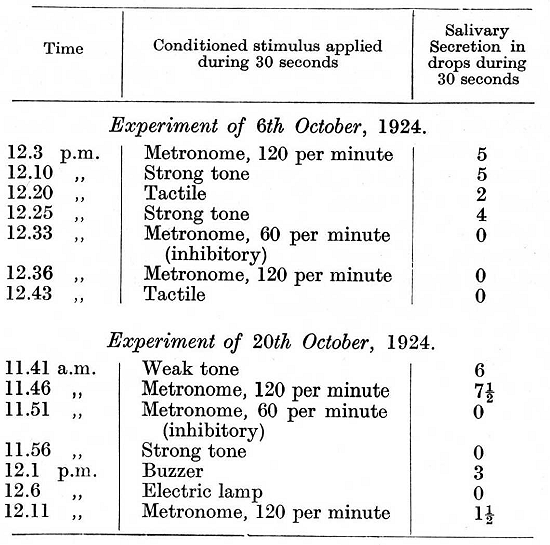
I have intentionally given several experiments with the initially positive reflexes in order to show how long and how persistently the normal relations were retained in spite of the disturbing influence of the metronome during the intervals between these particular experiments. On continuing the experiments further the general relation between the strength of the reflexes and the strength of the stimuli continued to be maintained in all experiments in which the metronome was not used. In those experiments, however, in which either of the rates of the metronome -- which themselves gave a secretion varying from 1/2 to 7 1/2 drops - was used there was invariably a disturbance of all conditioned reflexes following within the given experiment -- a disturbance in the form of complete inhibition or of one of its intermediate stages. It is of interest that the metronome [p. 306] rate of 120 per minute often produced a greater disturbance than the formerly inhibitory rate of 60. The experiments given below are taken from this period of experimentation.
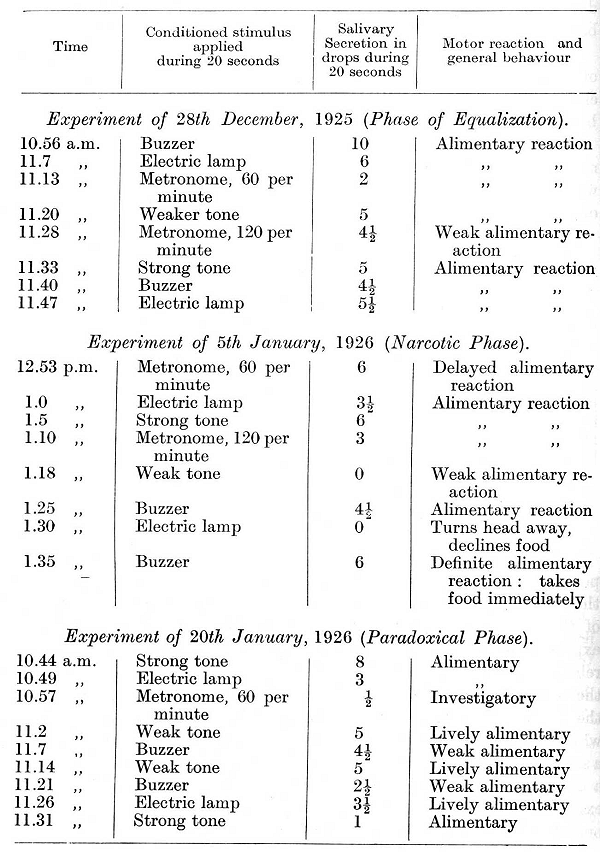
[p. 307]
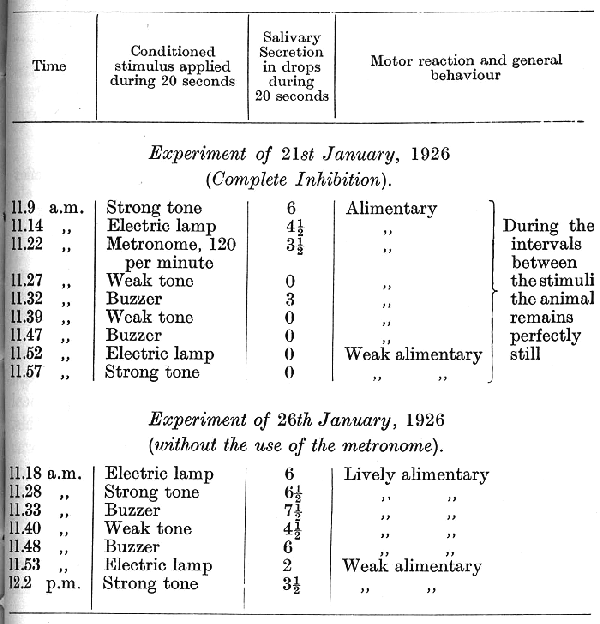
In the last experiment, though the relative strength of the reflexes remained unchanged, a general diminution in the strength of all the reflexes became apparent towards the end of the experiment. In view of this tendency all the stimuli were for several days reinforced after a very much shorter delay and the use of the metronome was discontinued. Subsequently to this, the delay was extended again, but only to 15 seconds instead of to 20 seconds as previously. Moreover, a new conditioned reflex to a bubbling sound was established; the stimulus belonged in this dog to the group of strong stimuli. As a result of this treatment the reflexes increased in strength and now showed no diminution towards the end of an experiment. After eleven days of such experimentation the rate of 120 beats per minute of the metronome was again tried: [p. 308]
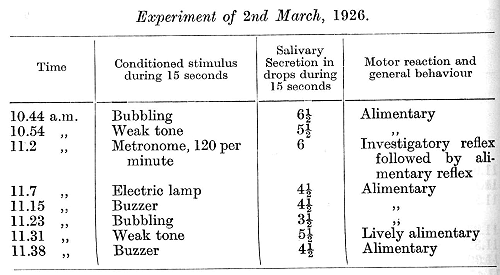
It was seen that the administration of the metronome led to an immediate disturbance of all the succeeding conditioned reflexes, which entered into a phase of equalization passing into the paradoxical phase. The disturbance, however, went much deeper than this. On the following day and for a long time afterwards the cortex was unable to withstand any kind of strong stimulus without undergoing complete inhibition. The fact that the maximum disturbance in the central nervous activity does not appear immediately on administration of the causative stimulus, but after one or more days has been observed in many animals. The following is an experiment which was performed on the day following the last administration of the metronome:
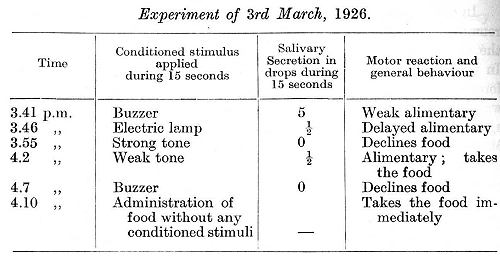
[p. 309]
To all appearance the dog was at this time perfectly healthy.
The cortical disturbance continued for 11 days without showing signs of any improvement, and we therefore determined to avoid any further use of the strong tone and to damp down the sounds of buzzer and bubbling. The following is the first experiment:
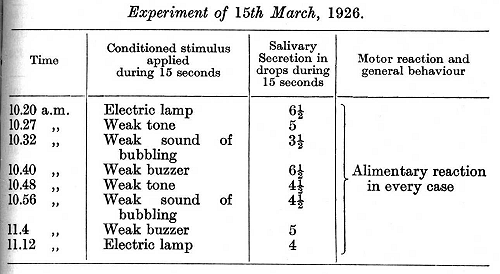
The experiments gave similar results on nine successive days. After this the strong stimuli were again applied, with the following result:

[p. 310]
When free upon the floor the animal behaved quite normally and took the food with avidity. After an interval of one day an experiment was performed again with the use of only weak stimuli, and all the reflexes were found to be present.
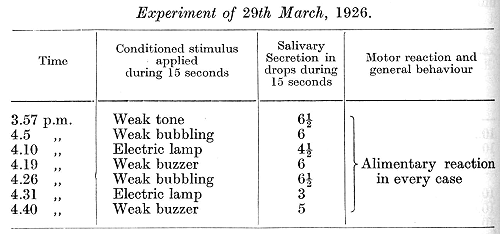
To sum up, these experiments show that the transformation of an inhibitory point of the acoustic analyser into an excitatory one occurred only gradually and imperfectly. Moreover, and this is more important, it rendered this point abnormal so that its stimulation by the conditioned stimulus of the metronome immediately led to a profound disturbance in the activity of the entire cortex, leading finally to an inability to withstand any strong conditioned stimuli without passing into different phases of inhibition, including the phase of complete inhibition. At first the normal activity of the cortex was comparatively quickly restored on discontinuing all stimulation of the abnormal point. At a later period, with further stimulation of this point, the disturbance took on a more permanent character. Since other auditory stimuli continued at this time to act quite normally the disturbance must be regarded as a result of a strictly localized functional interference in the acoustic analyser, a chronic functional lesion of some circumscribed part, the stimulation of which produces an immediate effect upon the function of the whole cortex, and finally leads to a protracted pathological state.
These observations to my mind reveal once more in an almost tangible form the mosaic character of the cortical activities which has been discussed already.
The disturbance in the activity of the entire cortex which has just been described can be regarded as being produced in either of [p. 311] two ways. First, it is possible that the excitation evoked by stimuli acting upon the area of cortical disturbance now rapidly passes over inhibition. Such a transition into inhibition is at first restricted to the immediate area around the original point of cortical disturbance but ultimately irradiates to involve the whole of the cortex. Second, it is possible that the stimuli act upon the area of cortical disturbance as injurious agents, so that, exactly as in the case of any injurious agents acting on any other part of the body, the entire cortical activity becomes inhibited on account of external inhibition.
In either case it is obvious that the localized disturbance of the acoustic analyser is again the result of a clash between excitation inhibition.
Besides those cases which have been related in the present and in the preceding lectures many others have been observed in which a similar clashing of the two antagonistic nervous processes led sometimes to a temporary, sometimes to a more prolonged disturbance of the normal activity of the cortex, in the form of a lasting predominance of one or the other process. In many cases these pathological disturbances could not be remedied by any of the measures which were applied. These abnormal conditions developed either during the establishment of very difficult differentiations -- especially in the case of successive compound stimuli [experiments of Dr. Ivanov-Smolensky, Dr. Eurman and Dr. Zimkina] -- or on immediate transition from inhibitory cutaneous stimuli to excitatory ones -- especially when the differentiation depended upon a definite rate of stimulation of one and the same place on the skin. When in the latter case the experiments were performed on an excitable and aggressive type of animal [experiments of Dr. Federov] the general excitation reached such an intensity that it became impossible to continue the experiments. The animal was, however, cured by prolonged administration of bromides and by disuse of both positive and negative tactile stimuli. In a dog of more inhibitable type [experiments of Dr. Petrova] there developed under similar conditions an abnormal focus, to all appearance strictly localized, in the cutaneous analyser, just as in the experiments of Dr. Rickman with the acoustic analyser. The positive stimulation of this point invariably led to a diffuse spreading of inhibition which persisted throughout the given experiment or even during a few subsequent days. Further experiments with this dog were prevented on account of its falling ill of nephritis. [p. 312]
With regard to the pathology of the cortex, there remain to be mentioned another set of cases in which the disturbances verge almost impreceptibly [sic] into the normal. These cases are the result of a permanent congenital weakness of the nervous system of the animal, which under definite conditions is rendered abnormal, while a more resistant nervous system remains normal. However, before, describing these it is necessary, for the sake of clearness, to discuss briefly those external stimuli which directly lead to inhibition of the cortical elements. These are of three kinds--monotonously recurring weak stimuli, very strong stimuli and unusual stimuli. All these can result either from the appearance of some new stimulus or from a new grouping of old stimuli. The conditions of our lives as well as of the lives of animals provide many occasions for the action of such stimuli, and it is not necessary to give any special examples. The biological significance of the development of cortical inhibition in response to such stimuli can easily be perceived. If stimuli of considerable strength, and especially continually changing stimuli, determine -- as they must do in order to maintain the delicate equilibration of the organism in its surroundings -- an alert state of the cortex, then it is quite reasonable to suppose that weak and monotonous stimuli which do not call forth any activity should tend towards inhibition so as to rest the cortical elements and give them time for recovery after a preceding activity. The inhibitory influence of very strong stimuli can be regarded as a reflex of "passive self-defence," as, for instance, in the case of hypnosis. The immobility of the animal makes it less noticeable to the enemy, and thus abolishes or diminishes the aggressive reaction of the enemy. It has been seen that the presence of something unusual also leads to a limitation of movement, which again may possess "survival value" for the animal, since in the new conditions its usual reactions might not be appropriate and might lead to some injury. Of course in the case of a new, even small, change in the environment, two reflexes appear -- a positive one, the investigatory reflex, and a negative one, which might be described as a reflex of caution and restraint. Whether these two reflexes are independent or whether the second is a sequence to the first and results from external inhibition or negative induction cannot be settled at present. The second supposition seems to me the more probable. The mechanism by which these three different types of stimuli bring about inhibition will be discussed in a further lecture. [p. 313]
A big flood which occurred in Petrograd on the 23rd September, 1924 afforded us an opportunity to observe in our dogs prolonged neuro-pathological disturbances which developed as a result of the extremely strong and unusual external stimuli consequent on the flood. The kennels of the animals which stood on the ground at a quarter of a mile's distance from the main building of the laboratory were flooded with water. During the terrific storm, the breaking of the waves of the increasing water against the walls of the building and the noise of breaking and falling trees, the animals had to be quickly transferred by making them swim in groups from the kennels into the laboratory, where they were kept on the first floor, all huddled up together indiscriminately. All this produced a very strong and obvious inhibition in all the animals, since there was no fighting or quarrelling among them whatever, otherwise a usual occurrence when the dogs are kept together. After this experience some of the dogs on their return to the kennels showed no disturbance in their conditioned reflexes. Other dogs -- those of the inhibitable type -- suffered a functional disturbance of the cortical activities for a very considerable period of time, as could be disclosed by experiments upon their conditioned reflexes.
One of the dogs has already been mentioned in these lectures [experiments by Dr. Speransky -- a strong healthy animal, but very easily subjected to inhibition, with all conditioned reflexes normally of considerable magnitude, very constant and very precise so long as the environing conditions were kept rigidly constant. This dog had ten alimentary conditioned reflexes, six positive and four negative (differentiations). Out of the positive reflexes three were auditory and three visual. The buzzer, which was the strongest of the auditory stimuli, evoked the largest secretion. The three visual reflexes were equal in their secretory effects and smaller by about one-third than the auditory. A week after the flood the dog was brought in to the experimental room and placed in its stand. The animal was abnormally restless and all conditioned reflexes were practically absent, and, though usually very ready for food, the animal now would not touch the food and even turned its head away. During three days while the animal was purposely left without food its general behaviour during the experiments remained unaltered. On considering various possible interpretations we reached the conclusion that this extraordinary behaviour of the animal must still be an [p. 314] after-effect of the flood, and the following method of combating the disturbance was adopted: Instead of leaving the animal alone during the experiment the experimenter now remained in the same room with it, while I myself conducted the experiment from the outside. All the reflexes showed an immediate restoration in the first experiment and the animal took the food with avidity, but it was sufficient for the experimenter to leave the animal alone for all the abnormal symptoms to recur. In order to re-establish the reflexes permanently it was necessary to adopt the above course, of alternately leaving and entering the room, for a considerable period of time. On the eleventh day of this treatment a conditioned stimulus which had not been employed since before the flood was now for the first time again employed, namely, the buzzer, which was the strongest stimulus, and which before the hood constantly evoked the largest secretion. After the application of the buzzer all the remaining conditioned reflexes almost completely disappeared, the animal again declined the food, became very restless and continuously stared at the door. Under the influence of the same special stimulus, namely, the presence of the experimenter in the room with the animal, the reflexes were again gradually restored, but repetition of the buzzer after an interval of five days produced the disturbance afresh. The buzzer was then applied only when the experimenter was in the room with the animal, but even so, normal relations only returned gradually and very slowly. On many occasions a phase of equalization of the reflexes was observed after administration of the buzzer, the reflexes often diminishing and the animal declining the food. Perfectly normal reflexes were at last obtained after forty-seven days of experimentation, i.e. two months after the flood. We now made the following experiment. A small stream of water was allowed to trickle silently beneath the door into the animal's room. The water formed a small pool on the floor next to the table on which the dog stood in its stand. This experiment, which is represented in full on page 315, was conducted in the absence of the experimenter from the animal's room.
Several months after, when the reflexes were perfectly normal and the buzzer had intentionally not been used for a considerable length of time, a first fresh application of the buzzer gave a reflex which was of greater intensity than the reflexes to the other stimuli, but on repeating the stimulus of the buzzer once every day for several days the secretory effect gradually diminished, and finally [p. 315] the buzzer became not only entirely ineffective of itself, but its applications now led also to a diminution of all the other reflexes. It is of interest that at this stage of the experiment the presence not

Evidently under the effect of an extremely powerful and unusual stimulus the cortical cells, which in this dog already had a tendency to inhibition, now permanently became still more susceptible to inhibition. Stimuli to which the dog had already for a considerable time been indifferent (the general environment of the experiment), and also strong agencies which had acted before as powerful conditioned stimuli (buzzer), produced a strong inhibitory effect upon the cortical elements, the resistance of which was now diminished. Minutest components of the extraordinary stimulus of the hood were sufficient to evoke the same abnormal reaction.
The dog upon which the experiments described in detail at the beginning of the present lecture were performed had also passed through the experience of the flood, with resulting disturbances, which, though based on a similar predisposition to inhibition, assumed a somewhat different character from those observed in the first dog. Two experiments are given below, the one performed the day before the flood (22nd September, 1924), the other on the third day after the flood (26th September, 1924).
(The experiments by Dr. Rickman upon the same dog, given in the first part of this lecture, were performed, it will be remembered, much later than this, December 1925 -- March 1926.)
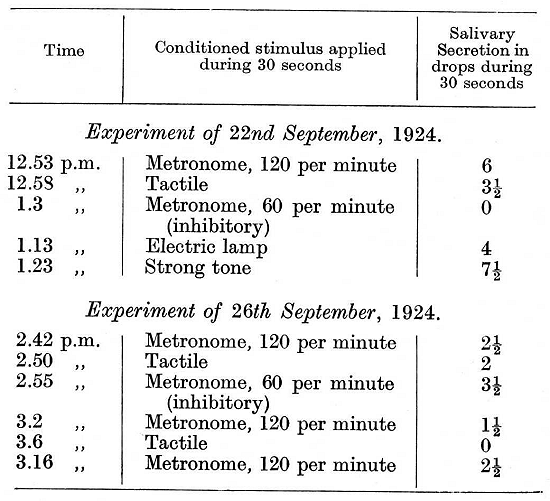
In the experiment of the 26th September the animal accepted the food, but the positive conditioned reflexes were diminished and the maximum secretory effect was obtained by the negative stimulus (2.55 p.m.-ultra-paradoxical phase). Experiments were now performed in which for a very long time the inhibitory rate of the metronome was not used. The positive reflexes were satisfactory and approached the normal, but even now on a single fresh application of the inhibitory stimulus, in any experiment, all conditioned reflexes were greatly diminished or even abolished, not only in the given experiment but during several succeeding days. The following are two examples:
During the period of recovery of the conditioned reflexes when the inhibitory stimulus was not used all the different phases of transition between complete inhibition and the normal positive effect were observed. In the beginning the recovery was favoured by the usual method of interrupting the experiments for a few days and shortening the isolated action of ah the conditioned stimuli. Soon, however, these methods became of no further use. The first [p. 318] one or two reflexes in the beginning of an experiment were generally very weak, and all the rest failed altogether. The animal grew lethargic and persistently declined the food. As a final measure the experiments were conducted with the animal kept free on the floor instead of in its stand. The beneficial effect of this method lies partly in the removal of the inhibitory influence of the stand, and partly in the introduction of additional excitatory impulses from the muscles and joints. The measure proved efficacious. The reflexes began gradually to return and progressively gained in strength. The animal accepted the food and normal relations became finally re-established. The administration of the negative stimulus led during the first seven days to a disappearance of the reflexes for the rest of that particular experiment. This inhibitory effect was not so apparent at the beginning of the experiment on the following day. In the course of the succeeding two weeks the prolonged inhibitory after-effect gradually disappeared and the inhibitory stimulus could now be practised during a single experiment more often. The differentiated inhibitory stimulus was repeated several times in every experiment, and concentration of the inhibition was accelerated by the immediate application of the corresponding positive stimulus. But it was only after two months of experimentation with the animal on the floor, and eight months after the flood, that it was found possible to return to the usual experiments with the animal in the stand.
It is thus seen that the powerful and unusual stimuli arising from the flood increased the susceptibility of the cortical elements to inhibition to so great an extent that even a comparatively minute intensification of inhibition from the outside, in the form of a conditioned inhibitory stimulus, rendered impossible for a long time any existence of positive conditioned reflexes under ordinary experimental conditions.
All these experiments clearly bring out the fact that a development of a chronic pathological state of the hemispheres can occur from one or other of two causes: first a conflict between excitation and inhibition which the cortex finds itself unable to resolve; second the action of extremely powerful and unusual stimuli.
I have still to describe the pathological state of another animal [experiments of Dr. Vishnevsky], but unfortunately I cannot state definitely whether its present state depends only upon a congenital defect which has been accentuated by its general conditions of life [p. 319]-- age, pregnancy and so on -- or whether it was produced as a result of the hood, as in the case of the two preceding animals. This animal was described in the preceding lecture (p. 287) as belonging to an extremely inhibitable type. Previously to the flood it had been left for a long time without observation, and it was not until four months after the flood that experiments with the dog were resumed. It has already been mentioned that a long time before the hood this dog served for a considerable number of valuable experiments. Now, in spite of all the therapeutic measures which have been applied, the animal still cannot be employed for experiments upon our usual problems. All that I can do, therefore, in the present lecture is to describe its condition. The scope of the normal life of the animal, at any rate under laboratory conditions, has narrowed down considerably. In the laboratory it reacts to the minutest stimuli either by a passive defence reaction (an investigatory reflex which is immediately followed by inhibition of all movements and even refusal of food) or, as an exception to its type, it sleeps. Only in two ways can any return to normal conditions of life be brought about in this dog: either there must be adopted a quick transition from the conditioned stimulus to its reinforcement with food (within 1-2 seconds from its beginning), or else the experiments must be conducted with the animal on the floor and with the experimenter constantly moving about in the same room. In the latter case the animal trots after the experimenter during the whole of the experiment, but even so the reinforcement of the conditioned stimulus must not be too much delayed. Both methods effect improvement in the same manner, the animal now ceasing to react to small changes in the environment in the same way that it did before, so that it is now able to take the food unless disturbed by stimuli of unusual strength. Conditioned reflexes begin, under these conditions, to return. It is, however, only necessary to delay reinforcement until 5-10 seconds from the beginning of the conditioned stimulus for the animal quickly to grow drowsy and even fall asleep over its plate while taking the food. This truly extraordinary state of the nervous system may be pictured as a state of extreme exhaustion -- a perfect example of the so-called faiblesse irritable. The disturbance obviously has its seat in the cortical elements, since the delicate reactions of the nervous analysers are essentially an intrinsic function of the cerebral cortex. A more detailed investigation of this dog is being conducted at the present time.