(Return to index)
Posted March 2001
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE XIX
Pathological disturbances of the
cortex, result of surgical interference:
(a) General disturbances of the
cortical activity. (b) Disturbances of the acoustic analyser.
Having attained, on entirely objective lines, to a measure of understanding concerning at least the main aspects of the physiological activity of the cortex, we naturally became interested to apply the method of conditioned reflexes to the study of cortical localization of functions in an endeavour to determine the importance of different parts of the brain for the normal functioning of the cortex as a whole. Experiments were made in this direction even in the early stages of our research (see my communication at the International Medical Congress at Madrid, 1903). The only method so far available for such a study consists in observing the effects of partial destruction or complete extirpation of different parts of the cortex. This method naturally suffers from fundamental disadvantages, since it involves the roughest forms of mechanical interference and the crude dismembering of an organ of a most exquisite structure and function. Imagine that we have to penetrate into the activity of an incomparably simpler machine fashioned by human hands, and that for this purpose, not knowing its different parts, instead of carefully dismantling the machine we take a saw and cut away one or another fraction of it, hoping to obtain an exact knowledge of its mechanical working! The method usually applied to the study of the hemispheres or other parts of the central nervous system is essentially as primitive as this. Hammer and chisel, the saw and the drill; these are the instruments which must be used to open up the strong protective skull. Then we tear through the several layers of enveloping protective membranes, rupturing many blood-vessels, and finally, we injure or destroy whole lumps of the delicate nervous tissue in different mechanical ways -- concussion, pressure and incision. But such is the marvellous functional resistance and the peculiar vitality of the living substance, that, in spite of all these gross manipulations, within the lapse of only a single day it is sometimes impossible without special and exact investigations to observe [p. 321] anything abnormal in animals submitted to cerebral operations. Accordingly, even by these primitive methods, some insight into the functions of the cortex can be gained. But the obvious usefulness of these crude methods should on no account satisfy the physiologist. He should strive to apply new advances of technical science and to seek ever new and more appropriate methods for the study of the exquisite mechanism of the hemispheres. Naturally the methods available for the investigation of the cortex at present, by means of extirpation of different parts, can but lead to entangled pathological states, and even the most guarded deductions with regard to the constitution of the cortex cannot therefore be ensured against a high probability of error. Indeed, since the special function of the cortex is to establish new nervous connections and so to ensure a perfect functional correlation between the organism and its environment, every disturbance of any part of it will be reflected upon the whole mechanism. Besides this direct influence of the operative procedure, which may reasonably be expected with time to diminish spontaneously, there is another very serious complication of the operation which appears later on -- namely, the development of a scar at the place of cerebral lesion which now becomes a source of irritation and leads to further destruction of the surrounding parts. On the one hand the scar, owing to its mechanical irritation of the surrounding normal parts of the brain, sets up recurrent outbursts of nervous excitation; on the other hand, owing to pressure, distortion and rupture, it progressively disintegrates the hemispheres. I have been unfortunate in attempting to improve the operative technique, having made, as I now think, a big mistake. In order to obviate haemorrhage during the operation I used to remove in the dogs, long before the operation on the brain, the temporal muscles which cover the skull; this resulted in a partial atrophy of the bones of the skull, so that these could now be opened, often without the loss of a single drop of blood. But the dura mater in these cases also undergoes considerable atrophy, becomes dry and brittle, so that in most cases it is impossible to make use of it to close up the cerebral wound completely. As a result the wound was left after the operation in direct communication with the more external tissues, which led to the formation of a very hard scar ultimately penetrating and growing into the cerebral tissue. Almost every animal that was operated upon suffered from attacks of convulsions which on some occasions occurred so soon as five to six [p. 322] weeks after the operation. A few animals died during the first attack, but more usually the convulsions were not severe in the early stages and occurred at infrequent intervals. In the course of several months they recurred more frequently and increased in force, finally either proving fatal or else leading to a new and very profound disturbance of the cortical activity. Therapeutic measures in the form of repeated anaesthesia or extirpation of the scar were found to be unreliable, though sometimes they were unquestionably effective.
Besides the difficulties arising on account of the purely surgical interference itself, the experimenter has to take into serious account a further difficulty which is especially pronounced in the case of the cerebral hemispheres. In the living organism we constantly discover different mechanisms by means of which functional compensation of damaged and destroyed parts is effected. In the nervous system such vicariation and compensation of function occur in the most extreme degree. It is well known in the spinal cord -- where it is facilitated by the complicated and varied course of the nerve fibres -- and also in the peripheral nervous system. The effect of mechanical destruction of localized parts is thus neutralized to a considerable extent by compensation. In the highest parts of the central nervous system, which regulate the major part of the internal and external activity of the organism, the principle of compensation and vicariation must be still more pronounced.
Having at our disposal the method of conditioned reflexes, related to different individual analysers, we attempted to determine and study the pathological disturbances affecting the entire cortex following the extirpation of one or another part, and also to use this study so far as possible in investigating the problem of the general construction of the hemispheres and the significance of its separate parts.
The first change which follows the
extirpation of some part of the cortex is the almost invariable disappearance
of conditioned reflexes; but in the majority of cases it is only the "artificial"
conditioned reflexes which disappear, i.e. those which were established
in the laboratory, being therefore relatively recent and little practised.
If the "natural" conditioned reflexes have also disappeared they are always
the first to reappear; but usually no disappearance of the natural conditioned
reflexes could be observed even though tested immediately after the recovery
from the anaesthetic ad- [p.323]ministered during the operation. Examples
of the greater resistance of conditioned reflexes to "natural" stimuli
as compared with those established to "artificial" stimuli occur in every
research upon partial extirpation of the cortex and there is therefore
no need to give any special examples. As a rule the conditioned reflexes
disappear after the operation, whether it is performed on one or both of
the hemispheres and on whatever portion of them it is carried out. The
absence of "artificial" reflexes persists for different lengths of time,
varying from a single day to several months.
Usually the greater the lesion the more prolonged is the absence of the reflexes; but considering the whole number of our experimental animals we find in this respect not a few exceptions. Even in animals in which the operations were apparently identical as to the place and extent of the lesion, there are great variations as to the length of time during which conditioned reflexes are absent. It is highly probable that, apart from the skill and neatness with which the operation was performed, the ultimate extent of the irritation and the destruction of the tissue depend upon the anatomical and functional peculiarities of the operated animal. After the operation the conditioned reflexes never return all at once, but in sequence, [p. 324] depending not only, as already mentioned, upon the stability of the reflex, but also upon the locality of the lesion. Generally speaking, reflexes belonging to those analysers which are most distant from the place of the lesion recover quickest. For example, after the removal of the gyrus pyriformis [experiments of Dr. Zavadskyl the conditioned reflex from the surface of the mouth (the "water reflex" which will be discussed later) reappeared on the eleventh day, the reflex to the smell of camphor on the eighteenth day, the reflex to an increase in the general illumination of the room on the twenty-fifth day, while the reflexes to conditioned tactile stimuli failed to recover even by the thirty-fifth day. The experiments demonstrate a certain disturbing influence which has spread from the point of lesion over the mass of the hemispheres and has then receded to the point of origin. This disturbing influence is, of course, generally speaking, due to the irritation caused by the lesion. It has already been shown that the influence of very strong stimuli, and even a conflict between nervous processes of opposite sign, lead to a prolonged inhibitory after-effect. It is, therefore, only natural to expect the same to result from a surgical destruction of a portion of the cerebral cortex itself.
When the conditioned reflexes finally get re-established they are found not only to regain their normal strength but often to exceed it, often also becoming considerably more stable than before. The inhibitory process, on the other hand, grows weaker. Many examples of this can be quoted from the experiments at our disposal. Thus, after extirpation of a part of the acoustic area of H. Munk in two dogs (see Fig. 13), the conditioned alimentary reflexes not only made a complete recovery but considerably increased in strength, and kept constant throughout every single experiment, whereas before the operation they used to decrease considerably towards the end of an experiment [experiments of Dr. Eliason]. The increase in the strength of the conditioned reflexes was still more conspicuous in another dog, after removal of the occipital lobes. The alimentary reflexes in this dog measured before the operation 1-2 drops during the isolated action of the conditioned stimuli; after the operation 13 drops [experiments of Dr. Koudrin]. In many dogs there is observed after an operation a very definite prolongation of the salivary secretion which follows the administration of an unconditioned stimulus: the length of time required for complete extinction of the reflex also becomes in many dogs very prolonged: the develop- [p. 325]ment of differentiations and conditioned inhibitions becomes more difficult and very often salivary secretion is observed in between the application of the stimuli -- this never happening before. This last is most probably to be accounted for by dis-inhibition of the reflex to environment (see the seventh lecture). It is an open question whether this weakening of the inhibitory effect is the result of an increase in the intensity of the excitatory process or whether the excitatory process is revealed on account of weakening of inhibition itself.
Another peculiarity with respect to inhibition is observed after surgical interferences. The inhibitory process becomes inert; and so to speak inflexible. As we saw before, in normal animals the inhibitory after-effect becomes with practice concentrated with regard to its duration as well as its extent. In the post-operative period this concentration proceeds extremely slowly and is imperfect. This inertia of the inhibitory process is observed not only in the reflexes belonging to the analyser which has been surgically damaged, but also in reflexes belonging to other analysers [experiments of Dr. Krasnogorsky].
In this manner the endeavour to demonstrate by means of experiments with extirpation the disappearance out of the normal cortical activity of the functions related to the extirpated part is complicated in the first period after operation by the general effect which the operation has upon the hemispheres as a whole. Although this unfortunate complication slowly and gradually disappears, it is followed, as already mentioned, by a second complication which also affects the hemispheres as a whole, and which depends on the development of soar tissue. The effect of the scar varies widely in different cases. After one and the same operation the injurious effect of the scar develops in some cases quickly and is severe; in other cases it is slow and feeble. Unfortunately the former was much the more common case in our experiments. The most usual effect of the scar consists in recurrent attacks of convulsions, sometimes affecting the whole body, and sometimes localized in one or another group of muscles. Such explosive outbursts of excitation in the hemispheres have a, very pronounced after-effect upon their activity. In this respect one should distinguish changes which follow weak and rare attacks from changes following violent attacks or frequently recurring attacks of moderate strength.
We shall start by considering the effect of the former. On [p. 326] observing conditioned reflexes from day to day it is very often possible to predict the approach of an attack of convulsions with accuracy. If, suddenly, without any obvious cause the conditioned reflexes diminish in strength and then disappear, it is an infallible sign of an approach of an attack of convulsions. Sometimes it is possible to notice a still earlier sign, consisting in the disappearance of differentiations, i.e. in a disturbance in the inhibitory process After an attack has passed, the re-establishment of the conditioned reflexes occupies a variable length of time, requiring sometimes hours, and sometimes days. In some instances the re-establishment of conditioned reflexes assumes a complicated character. Immediately after the termination of the attack conditioned reflexes are present, but, some time after, they again disappear, now for a considerable length of time. This may possibly be explained by an initial irradiation of the outburst of excitation, followed subsequently by concentration and negative induction. So far as the effect of very strong or frequent attacks is concerned their after-effect is very variable. In one dog they were apparently the cause of an absolute deafness; another dog, which after the operation behaved quite normally towards men and towards other dogs and food, began after a severe attack of convulsions to dodge and run away from other dogs and from men. Finally a fresh attack killed her. A third dog, after numerous and frequent attacks, showed quite peculiar symptoms. The dog will be described later in detail, but meanwhile I shall describe one of the symptoms. In this dog, after the attack, all the reflexes returned; they had, however, to be used with practically simultaneous reinforcement. Even small delays (up to 5 seconds) quickly led on repetition to disappearance of the conditioned effect, with refusal of food and development of sleep. Obviously the dog suffered from a chronic state of faiblesse irritable such as was described in reference to another dog at the end of the preceding lecture. After each recurrence of the attack this peculiar symptom became more pronounced. It is natural to regard this change as due to a functional exhaustion of the cortical elements consequent on the convulsions; the cortical elements now, under the action of external stimuli, rapidly undergo a transition into an inhibitory state, the connection of which with sleep has been discussed previously. In this dog the functional exhaustion, like the excitability producing it, affected the entire cortex. It is often, however, restricted to the particular analyser which was [p. 327] most directly effected by the surgical interference. Examples of this will be given later.
In some cases the effect of the scar manifests itself in a different manner, being limited, to a hyper-excitability of analysers other than the motor one, and therefore not associated with convulsions. One dog after extirpation of the frontal lobes [experiments of Dr. Babkin] quickly recovered from the effects of the operation, but two months later it developed an extreme cutaneous hyperaesthesia which lasted for ten days: the animal would howl at the most gentle touch and even at its own movements, and after this would shrink down into a heap on the floor. Evidently the scar in the cutaneous analyser served as a source of irritation to the cortical area of localization connected with the receptors of injury (subjectively pain), if indeed such analyser exists in an independent localized form.
A still more interesting case was presented by another dog after a partial extirpation of the cortical part of the cutaneous analyser [experiments of Dr. Erofeeva]. One and a half months after the operation a vigorous attack of convulsions occurred. During this attack the animal was subjected to a further operation, the scar, which gave outgrowths considerably beyond the site of the original lesion, being carefully removed. The convulsions did not recur after the operation, but another form of disturbance developed which lasted at each recurrence for several days. When either experimenter or food came into the field of vision of the left eye (the animal being operated on the right side) it quickly turned away, and if free ran away, showing signs of extreme excitation. The same stimuli when applied from the right side of the animal produced no abnormal reaction. Often, free and on its own, the dog would suddenly glance to the left, quickly jump up and run madly away. This can all be interpreted if we assume that some remaining portions of the scar directly irritated the visual analyser on one side, thereby producing a distortion of the effect of the external stimuli falling on the retina and altering the significance of the visual object, which assumed in the dog's cortex unusual and extraordinary aspects to which the animal reacted as to any concrete and definite stimulus -- exactly in the same manner as happens also with normal animals in response to any extraordinary visual stimulus. In short, the scar produced a phenomenon of illusion. Obviously a similar condition obtained in the dog previously mentioned, which after an attack of convulsions began to run away even from its usual attendant and from the food, [p. 328] exhibiting a violent general excitation. It is probable that the outburst of excitation resulting from the scar, after having freed the motor area of the cortex, was still retained for some time in the visual analyser. It is legitimate to regard these cases as the equivalent of epileptic disturbances of the motor analyser.
The foregoing observations led to our planning a detailed investigation of the effects of direct stimulation of the different cortical analysers. By permanently healing electrodes in different points of the hemispheres we are hoping to produce in our dogs by means of electrical stimulation definite changes in the reactions to our usual conditioned stimuli. The difficulties of technique have been overcome and the experiments are at present in progress.
It is unfortunate that the majority of our experiments with extirpation were performed in the earlier period of our research, when we had not definitely realized the peculiarities of the different individual types of the nervous system and had not yet any knowledge of the pathological effects arising under the influence of functional, i.e. non-surgical, disturbances.
After this preliminary review of the general effects of surgical interference with the cortex I shall describe in detail the results obtained by the fullest possible application of the method of conditioned reflexes to our operated animals with the object of determining the physiological significance of the entire cortex and of larger or smaller parts thereof.
In some of our animals a complete extirpation of the whole cortex was carried out (one animal survived the operation for 4 1/2 years). This operation had already first been introduced by Goltz, but it was undertaken in our experiments with the special object of determining by our method the relation of the cortex to the higher nervous activity of the dog [experiments of Dr. Zeliony]. The general behaviour of the animals after the complete extirpation of the cortex has been described in detail on several occasions by Dr. Zeliony himself. I shall therefore dwell only upon the relation of conditioned reflexes to the cortex. Since all the usual conditioned reflexes were definitely absent in these dogs after the extirpation, and neither old nor new reflexes could be obtained in spite of the most persistent reinforcing of artificial stimuli or testing for natural conditioned reflexes, we concentrated our attention upon one peculiar conditioned reflex, which, according to our previous experiments, in contrast to all other conditioned reflexes, was extremely stable. I refer to what [p. 329] we term "the water reflex " -- a conditioned reflex in response to stimulation of the receptor surface of the mouth. If, by means of an apparatus which is placed in the dog's mouth, water is injected after a few preliminary injections of acid, the water which under normal conditions evokes hardly any salivary secretion (at most 1-2 drops) now causes a copious secretion. Evidently the stimulation by liquid of some receptor nerve-endings of the mucous membrane of the mouth, coinciding with the effect of the acidity, acquires conditioned properties determining this large salivary secretion together with a corresponding motor reaction typical of the reflex to acid. The conditioned water-reflex, as will be shown later, possesses all the properties of a conditioned reflex. In the dog which survived the removal of the hemispheres the longest time (4 1/2 years), the water-reflex was established before the extirpation of the last portion of the hemispheres -- the extirpation being performed in stages. The reflex measured 8-10 drops for injection of 5 c.c. of water. Beginning from the sixth day after complete extirpation of the hemispheres 5 c.c. of 0.25% solution of hydrochloric acid were repeatedly injected practically every day, over 500 injections being made in all. Only after seven months of this procedure did a salivary secretion to water appear, which gradually increased in strength until it measured 13 drops for injection of 5 c.cs. of water. Was this, however, a conditioned reflex? Decidedly not. The reflex in this case differed fundamentally from the ordinary conditioned water-reflex. The most important difference was that it could not be made to undergo extinction, which occurs with extreme ease in the case of the real conditioned water-reflex in normal animals when water is injected several times in succession without the acid. In the decorticated animal the effect of repeated injection of water became constant. By observing the dog after the administration of water the true nature of this reflex was revealed. After injection of water the animal exhibited typical movements, which otherwise were observed only when the animal was hungry. It started walking to and fro with head bent low and twitching nostrils, as if reaching for something. On further investigation it became obvious that the contact of water with the mucous membrane of the mouth evoked in this dog a strong unconditioned alimentary reflex. This was corroborated by the fact that unconditioned reflexes, e.g. salivary reflexes, after removal of the cerebral cortex, become at first diminished, afterwards, however, gradually recovering their [p. 330] wonted strength and finally becoming very much increased above normal.
To present the final conclusion of these experiments with the utmost reserve, the cerebral cortex should be regarded as the essential organ for the maintenance and establishment of conditioned reflexes, possessing in this respect a function of nervous synthesis of a scope and exactness which is not found in any other part of the central nervous system.
Of the individual analysers most attention was paid to the acoustic, and I shall commence my description with this. An absolute deafness following extirpation of a part of the cerebral cortex was observed in three dogs. In two of these animals [experiments of
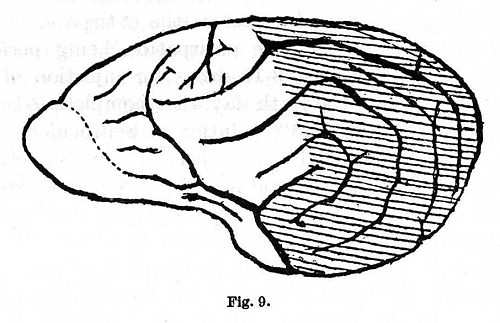
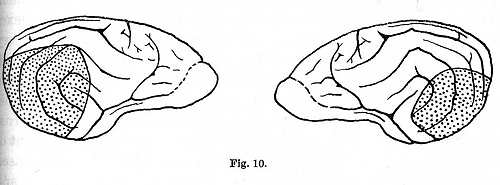
Dr. Koudrin] the cortical substance was removed posterior to a line starting from a point above and immediately behind the gyrus sigmoideus, stretching to the tip of the gyrus sylviaticus and then passing along the fissura fossae sylvii (Fig. 9). The operation was performed in two stages, first on one and then on the other side. Absolute deafness appeared immediately after the second operation. One of the dogs lived for nine months after the operation, the other for seven. In the third dog [experiments of Dr. Makovski] a bilateral extirpation was performed of the gyri sylviaticus posterior, ectosylvius posterior and suprasylvius posterior. On one side there were included also the middle and the frontal portions of these convolutions (Fig. 10). Absolute deafness occurred one and a half months after the operation. On the day previous to the deafness a weakening of the inhibitory processes was observed: this was probably followed during the night by an attack of convulsions. The animal lived after this, in apparently [p. 331] good health, for another month. During this period new conditioned reflexes were developed to stimuli belonging to the tactile, olfactory and visual analysers. The animal died during an attack of convulsions. Other animals, which were operated on in exactly the same manner as the last, continued to react to sound, though in some cases they survived the operation even longer.
How can we explain the absolute loss
of all auditory reactions? Since it must be regarded as definitely proved
that after complete removal of the whole cortex dogs still continue to
react to sound, it must be admitted that in the three cases of absolute
deafness described above there must have been some damage of sub-cortical
structures or a development of inhibition which spread into the sub-cortical
areas. The latter possibility ought not to be excluded,
since on histological examination (unfortunately rather crude) no damage of sub-cortical structures could be observed; further, in the first two dogs a general reaction to light, which at first entirely disappeared, returned two months after the operation, and a conditioned reflex to light even reached its pre-operative intensity; finally, it is well known how extremely easily different impulses originating within the hemispheres inhibit reflexes of lower centres. If such an inhibitory effect is accepted as the cause of deafness in the third dog, we have to assume that this inhibition spread solely throughout the ramifications of the acoustic analyser, without involving any other analysers.
As a rule the general auditory motor reactions (pricking up the ears and lifting the head), after removal of the temporal lobes or of the whole posterior half of both hemispheres, return within a few hours, or a day or two at most; in some cases they do not disappear at all. This general reaction to sound is an unconditioned investigatory reflex belonging to sub-cortical regions, since it remains [p. 332] present in dogs even after removal of the entire cerebral cortex. The rest of the auditory reactions must be attributed to the cortex, and these functions of the acoustic analyser at first entirely disappear after the limited operations described above, then return, never, however, recovering completely.
When, after bilateral removal of the temporal lobes, the investigatory reflex to sound has already returned, as well as conditioned reflexes to stimuli belonging to other analysers, all auditory conditioned reflexes are found still to be absent. Such a state may last for many days or even for several months, depending upon the extent of the operation. Moreover, it is of importance that both temporal lobes should be removed at one time, or in two operations shortly following one another, first on one side and then on the other. If the two operations are performed with a long interval between them the phase of complete disappearance of auditory conditioned reflexes may be absent. What does the temporary absence of auditory conditioned reflexes mean? Several possible interpretations can be advanced. First, it is possible that the cells of the acoustic analyser still remaining after the operation are rendered incapable of developing a state of excitation, and under the influence of external stimuli pass directly into an inhibitory state, it may be on account of being weakened by the operation, or on account of being decreased in number, or on account of being previously kept in reserve and not involved as a general rule in the activity of the acoustic analyser before the operation. Second, it is possible that after the operation the analysing function of the cortical part of the acoustic analyser is so diminished that all the sounds now affecting the dog, both inside and outside the laboratory, acquire identical qualities and therefore, more frequently than not, fail to coincide with the unconditioned stimulus, with the result that the conditioned significance of the definite acoustic stimulus disappears on account of extinction. Finally, it is possible that under the influence of the operation the synthetic activity of the acoustic analyser, involved in the maintenance or establishment of conditioned reflexes, itself weakens or temporarily disappears. Special experiments were performed in order to test these suppositions [Dr. Krijanovsky].
In one test use was made of an auditory
stimulus as a conditioned inhibitor. Two conditioned inhibitors of the
same alimentary reflex to camphor were established in a dog; one was a
tactile stimulus, the other auditory (tone d', 288 d.v. of a pneumatic
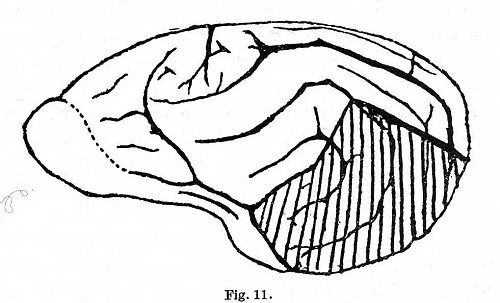
tuning fork). [p. 333] Three days after bilateral removal of the temporal
lobes (Fig. 11) the positive conditioned reflex to camphor had already
reappeared. The conditioned inhibitors remained practically without any
effect for several more days, but from the twelfth day on gave a full inhibition.
It was noticed now that any other sounds had exactly the same inhibitory
effect as the original conditioned inhibitor. All positive acoustic conditioned
reflexes were absent at this stage; neither the sound of splashing of acid
nor the sound of cracking of biscuits, to both of which under normal conditions
reflexes developed with extreme ease, could be made to evoke a conditioned
response. The fact that ally sound acted in the capacity of a real conditioned
inhibitor and not as an agent of external
inhibition was proved by means of destruction of the conditioned inhibition by reinforcing the action of the inhibitory combination by the administration of food and then re-establishing it by discontinuing the reinforcement. In control experiments the same destruction and re-establishment was repeated with the tactile conditioned inhibitor. It follows from these experiments that the sounds acted as true conditioned inhibitors, and, therefore, that the analyser was capable of performing an inhibitory function while the function of excitation was still absent. In other words, the analysing function of the cortex was impaired. Only several days after the above experiments did the positive conditioned reflexes to sound begin to reappear.
In a second method of testing our suppositions, in particular the supposition of extreme generalization of sounds, we used long-trace reflexes in which, as is known, the stimuli are generalized [p. 334] beyond the limits of a single analyser (Lecture VII). In the case of long-trace reflexes perfectly neutral stimuli assume the character of accessory conditioned stimuli, which act in the same manner as the original trace stimulus, i.e. the secretion begins after the same latent period. Now it was determined to test whether these conditioned accessory reflexes to auditory stimuli in general would still be present at the time of absence of the specific reflexes to these stimuli. For this purpose the dog had a definite trace reflex established to a tactile stimulus in which the pause between the end of the conditioned stimulus and the beginning of the unconditioned stimulus was two minutes. Usually the conditioned secretion started during the second minute of the pause. Ten days after complete removal of the temporal lobes the trace reflex to the tactile stimulus reappeared. On the twelfth day an auditory stimulus gave 8 drops during 4 minutes, the secretion starting during the third minute after the termination of the stimulus. On the seventeenth day the same auditory stimulus gave 38 drops during 6 minutes, the secretion starting as in the case of the primary trace stimulus during the second minute of the pause. It was only on the thirty-fifth day that the conditioned reflexes to the actual isolated action of the conditioned auditory stimuli first began to appear. This experiment shows once more that the auditory stimuli had assumed an extremely generalized character, so that sound as a general stimulus still continued to act, although individual sounds had lost their specific conditioned significance. It is, moreover, evident that the function of synthesis was not lost, and after these experiments special testing of the third supposition was unnecessary.
The two types of experiment just described probably belonged to two different post-operative states of the acoustic analyser, the first an earlier, the second a later stage. This is the more probable since in other dogs which were subjected to a similar operation (Fig. 12) we also observed a generalization of auditory conditioned simultaneous and short-delayed stimuli [experiments of Dr. Babkin]. The following is one of these experiments:
A conditioned alimentary reflex had
been established to a descending scale of four neighbouring tones, and
this was completely differentiated from the same scale taken in the ascending
direction. On the eighth day after removal of the temporal lobes the experiment
proceeded as follows: [p. 335]
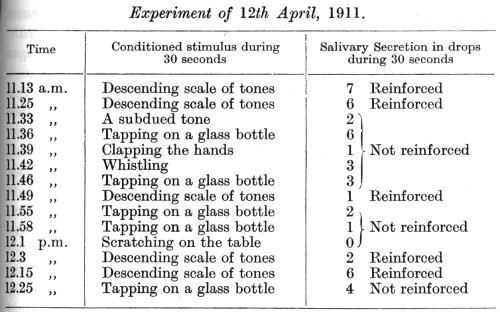
All the stimuli, except the scale of tones, used in the above experiment were " neutral," i.e. had never been reinforced.
It is seen from this experiment that besides the previously established auditory conditioned stimulus many other sounds, which never previously had any conditioned significance, now acted sometimes as effectively as the descending scale of tones. When these extraneous sounds weakened in their effect on account of non- reinforcement, the conditioned stimulus also considerably diminished in its effect (11.49; 12.3). On reinforcing the conditioned stimulus the effect of the other sounds also became increased (12.25 p.m.). The sound in its capacity of a conditioned stimulus had become extremely generalized, and its analysis had become impaired so as to be practically negligible. When the power of analysis begins to reappear the improvement proceeds very often with extreme slowness. First of all musical tones are distinguished from other kinds of sound, [p. 336] like knocks and noises. The differentiation between different tones remains imperfect for a very long time; in these experiments of Dr. Babkin, for example, differentiation between single tones only began to return through gradual stages two months after the operation.
The absence or the diminished precision of the analysing function of the acoustic nervous apparatus described is obviously identical with what H. Munk terms "psychic deafness." It is impossible, however, not to see the fundamental difference between the purely physiological and the psychological interpretation of these facts. According to the definition of Munk "the animal hears but fails to understand," and experimentation becomes sterile in the interpretation of "understand." But the physiological point of view opens up a vast field for experimentally investigating the different stages of the re-establishment of functions in the damaged acoustic analyser. Under normal conditions sounds are differentiated according to their strength, duration, continuous or interrupted character, point of origin, and nature -- whether tones, knocks, noises, etc. It must be expected -- and in this respect it can be stated that we have definite proof -- that in returning to normal the damaged acoustic analyser passes through different stages of activity, and only by detailed investigation of these stages can we hope ever to reach a complete understanding of the mechanism of acoustic analysis.
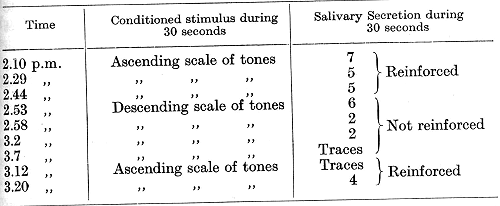
The foregoing does not complete our picture of the disturbances in the functions of the acoustic analyser after the removal of the temporal lobes. There is another, probably the most important, functional disturbance. It was noticed a long time ago, by many investigators, that after an operation upon the temporal lobes dogs cease to respond to their names. This was observed also in our experiments, and we believe that it can only be explained by a disappearance of the special analysis of conditioned auditory compound stimuli. In order to verify this point special experiments were performed by Dr. Babkin. Conditioned stimuli were established to different tones applied in different sequences or with different intervals between them. One definite sequence was used for a positive conditioned reflex, others for negative ones (differentiation). These differentiations, as mentioned in the eighth lecture, were much more difficult to establish than differentiations of single tones. Besides the differentiation of compound auditory stimuli, differentiations of single tones were also established. Both temporal lobes were then extirpated in these animals. The disturbances which followed the operation were [p. 337] exactly identical in the five dogs employed for the experiments. While the differentiation of single tones sooner or later became re-established with the same precision as before the operation (a differentiation of intervals of a single tone), there was never the slightest trace of any re-establishment of a differentiation of the successive compound stimuli, though most of the dogs were tested for 2-3 months after the operation, and one dog for nearly three years. In the latter dog [originally observed by Dr. Koudrin] the entire posterior part of the hemispheres was removed as in the two dogs previously described (see Pig. 9, p. 330). The final operation was performed on the 5th May, 1909. The experiments to be described were started towards the end of 1911. Alimentary conditioned reflexes were established to an ascending scale of tones of pneumatic tuning forks -- 290, 325, 370, and 413 d.v., and to a separate tone of a Stern's tone-variator -- 1200 d.v. The reflexes developed fairly quickly and a differentiation between the single tone of 1200 d.v. and a tone of 1066 d.v. was attempted through stages of differentiations from 600 and then 900 d.v. The final differentiation was successfully established. A differentiation of the descending scale, on the other hand, completely failed to be established in spite of 150 repetitions of the descending scale contrasted with 400 of the ascending scale. The reaction to calling the dog by name was also absent during the whole period of three years. The following example is taken from a late stage of these experiments (15th March, 1912).
To the same group of experiments probably belongs also the following, up to the present solitary, case [experiments of Dr. Eliason]. A conditioned alimentary reflex was established to a chord of tones of a harmonium 85-256-768 d.v. (F-c'-g"). When the [p. 338] reflex reached its maximum strength the different component tones were tried separately. They all produced a positive effect weaker than that of the chord but approximately equal in strength among themselves. The effect of intermediate tones was extremely small. After the removal of the anterior portions of the temporal lobes (Fig. 13) the relative effect of the different components underwent a considerable change. The effect of 768 d.v. and of the neighbouring tones disappeared altogether, though the reflex to the chord returned on the fifth day after the operation. The lower component of the chord -- 85 d.v. -- when tested alone began to act with increased vigour, its effect often being equal to that of the whole chord. What can be the explanation of the extremely definite results of these experiments? The first explanation that suggested itself was that
the reflexes to the higher musical tones had been selectively disrupted as a result of the operation. This, however, was absolutely disproved, since when the tone of 768 d.v. was reinforced independently of the chord it very quickly assumed independent and very definite properties as a strong conditioned stimulus. To our great regret this dog died before we could accomplish different modifications in the experiments. The absence of the effect of the higher tone before it was independently reinforced cannot be attributed to any difference in strength of the tones in the chord, since the highest tone was if anything stronger than the two lower ones. The observations are in accord with the theory of the existence of a special part of the acoustic analyser in which the synthesis and analysis of successive and simultaneous compound auditory stimuli is effected (the acoustic area of H. Munk). Such part of the acoustic analyser would provide a parallel with the undoubted projection of the retina upon a definite part of the visual analyser. According to this supposition such part of the acoustic analyser in the cortex must be regarded as a special receptive field which is connected with all the parts of the peripheral [p. 339] acoustic apparatus, so that on account of the specially favourable local structural peculiarities a facility is afforded for the formation of various and complicated connections, involving the establishment of reflexes to most complex compound auditory stimuli as well as their analysis. A partial destruction of this portion leads to a dropping out from the compounds of some of the individual components, and a complete destruction entirely eliminates the higher synthesis and analysis of compound stimuli. After complete removal of the temporal lobes auditory conditioned reflexes still continue to exist [Dr. Kalischer,1 and our own experiments], and an elementary differentiation can still be effected, while after extirpation of the whole cortex all conditioned reflexes entirely and permanently disappear. Only one conclusion, therefore, can be drawn, namely, that in the cortex, besides the special part of the acoustic analyser, there must exist some extensions of the analyser dispersed more widely over the cortex, and maybe throughout its whole mass. These elements owing to their dispersion are not able to enter into complex interconnections, though they can still perform an elementary synthesis and analysis. It is possible also that the simplification or limitation of activity of different parts of the acoustic analyser increases with their distance from the cortical "nucleus" of the analyser.
The hypothesis of such a distribution of the cortical part of the acoustic, and probably of any other, analyser seems to me to conform best with the available facts, and to open also an unlimited field for further investigation. It would fit in with the wide dispersion, which will be proved later, of any one analyser between the other analysers, far beyond the limits of the hitherto accepted localizations. It would also agree with the existence of a special "nucleus" in each analyser in which, on account of the density and the exceptional concentration of the elemental units of the given analyser, the higher synthetic and analytic activity is rendered possible. Again, it could also without difficulty explain the gradual improvement by practice of the activity of the remnants of the analysers, the functions of which are so limited immediately after the lesion of its nucleus. Ultimately it should also determine the limits to which such an improvement could extend. In the dog last mentioned the elementary [p.340] analysis of tones had probably reached the maximal degree possible, the loss of the higher synthesis and analysis remaining permanent even after three years of practice.
The hypothesis suggested, of such a wide distribution of analysers in the cortex, naturally sets up further problems for experimentation which should be used to test its validity. In regard to the acoustic analyser, before a partial extirpation there should be developed as great a number as possible of positive elementary and compound stimuli, and various differentiations. The different general properties of the elementary stimuli should be determined for each dog, their threshold strength, the conditions under which positive stimuli acquire an inhibitory character, the mobility of the inhibitory process and the degree of its after-effect, and so on. Only such further experiments can demonstrate definitely the changes which occur as the result of an operation, how far these changes affect the general properties of the reflexes and what changes constitute the direct result of damaging different parts of the analyser. During the period of re-establishment of the acoustic function after the operation special attention must be paid, as was previously mentioned, to the determination of the different transition stages. It is obvious that in order to carry out such a plan a healthy and prolonged existence of the animal after the operation must be ensured. Unfortunately this cannot up to the present be satisfactorily attained.
[1] O. Kalischer. "Zur Funktion des Schläfanlappens des Grosshirns." Sitzungsber, der Königl. preuss. Akademie der Wissensch. Physik. mathem. Kl, v. 10, p. 204. 1907.