
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE II
Technical methods employed in the objective investigation of the functions of the cerebral hemispheres. -- Response to signals as reflex action. -- Unconditioned and conditioned reflexes. -- Necessary conditions for the development of conditioned reflexes.
In the previous lecture I gave an account of the reasons which led us to adopt, for the investigation of the functions of the cerebral hemispheres, the purely objective method used for investigating the physiological activity of the lower parts of the nervous system. In this manner the investigation of the cerebral hemispheres is brought into line with the investigations conducted in other branches of natural science, and their activities are studied as purely physiological facts, without any need to resort to fantastic speculations as to the existence of any possible subjective state in the animal which may be conjectured on analogy with ourselves. From this point of view the whole nervous activity of the animal must be regarded as based firstly on inborn reflexes. These are regular causal connections between certain definite external stimuli acting on the organism and its necessary reflex reactions. Such inborn reflexes are comparatively few in number, and the stimuli setting them in action act close up, being as a rule the general physical and chemical properties of the common agencies which affect the organism. The inborn reflexes by themselves are inadequate to ensure the continued existence of the organism, especially of the more highly organized animals, which, when deprived of their highest nervous activity, are permanently disabled, and if left to themselves, although retaining all their inborn reflexes, soon cease to exist. The complex conditions of everyday existence require a much more detailed and specialized correlation between the animal and its environment than is afforded by the inborn reflexes alone. This more precise correlation can be established only through the medium of the cerebral hemispheres; and we have found that a great number of all sorts of stimuli always act through the medium of the hemispheres as temporary and interchangeable signals for the comparatively small number of agencies [p. 17] of a general character which determine the inborn reflexes, and that this is the only means by which a most delicate adjustment of the organism to the environment can be established. To this function of the hemispheres we gave the name of " signalization."
Before passing on to describe the results of our investigation it is necessary to give some account of the purely technical side of the methods employed, and to describe the general way in which the signalizing activity of the hemispheres can be studied. It is obvious that the reflex activity of any effector organ can be chosen for the purpose of this investigation, since signalling stimuli can get linked up with any of the inborn reflexes. But, as was mentioned in the first lecture, the starting point for the present investigation was determined in particular by the study of two reflexes -- the food or "alimentary" reflex, and the "defence" reflex in its mildest form, as observed when a rejectable substance finds its way into the mouth of the animal. As it turned out, these two reflexes proved a fortunate choice in many ways. Indeed, while any strong defence reflex, e.g. against such a stimulus as a powerful electric current, makes the animal extremely restless and excited; and while the sexual reflexes require a special environment -- to say nothing of their periodic character and their dependence upon age -- the alimentary reflex and the mild defence reflex to rejectable substances are normal everyday occurrences.
It is essential to realize that each of these two reflexes -- the alimentary reflex and the mild defence reflex to rejectable substances -consists of two distinct components, a motor and a secretory. Firstly the animal exhibits a reflex activity directed towards getting hold of the food and eating it or, in the case of rejectable substances towards getting rid of them out of the mouth; and secondly, in both cases an immediate secretion of saliva occurs, in the case of food, to start the physical and chemical processes of digestion and, in the case of rejectable substances, to wash them out of the mouth. We confined our experiments almost entirely to the secretory component of the reflex: the allied motor reactions were taken into account only where there were special reasons. The secretory reflex presents many important advantages for our purpose. It allows of an extremely accurate measurement of the intensity of reflex activity, since either the number of drops in a given time may be counted or else the saliva may be caused to displace a coloured fluid in a horizontally placed graduated glass tube. It would be much [p. 18] more difficult to obtain the same accuracy of measurement for any motor reflex, especially for such complex motor reactions as accompany reflexes to food or to rejectable substances. Even by using most delicate instruments we should never be able to reach such precision in measuring the intensity of the motor component of the reflexes as can easily be attained with the secretory component. Again, a very important point in favour of the secretory reflexes is the much smaller tendency to interpret them in an anthropomorphic fashion -- i.e. in terms of subjective analogy. Although this seems a trivial consideration from our present standpoint, it was of importance in the earlier stages of our investigation and did undoubtedly influence our choice.
For the purpose of registering the intensity of the salivary reflex all the dogs employed in the experiments are subjected to a preliminary minor operation, which consists in the transplantation of the opening of the salivary duct from its natural place on the mucous membrane of the mouth to the outside skin. For this purpose the terminal portion of the salivary duct is dissected and freed from the surrounding tissue, and the duct, together with a small portion of the mucous membrane surrounding its natural opening, is carried through a suitable incision, to the outside of the cheek in the case of the parotid gland, or under the chin in the case of the submaxillary gland. In this new position the duct is fixed by a few stitches which are removed when the wound has healed. As a result of the operation the saliva now flows to the outside, on to the cheek or chin of the animal, instead of into the mouth, so that the measurement of the secretory activity of the gland is greatly facilitated. It is only necessary for this purpose to adjust a small glass funnel over the opening of thc duct on to the skin, and for this we find a special cement prepared according to a formula of Mendeléeff[1] most useful. As an alternative, very suitable and accurate as s recording apparatus is a hemispherical bulb which also can be hermetically sealed on to the skin. From the bulb project two tubes, one pointing up and the other pointing down. The latter tube is used for drawing off the saliva which collects during each observation, while the former tube connects by air transmission with a horizontal graduated glass tube filled with coloured fluid. As the saliva flows into the hemispherical bulb the coloured fluid is displaced along the graduated tube, where [p. 19] the amount of secretion can be read off accurately. Further, it is not difficult to fix up an automatic electrically-recording device which will split up the displaced fluid into drops of exactly equal volume and reduce any lag in the movement of the fluid to a minimum.[2]
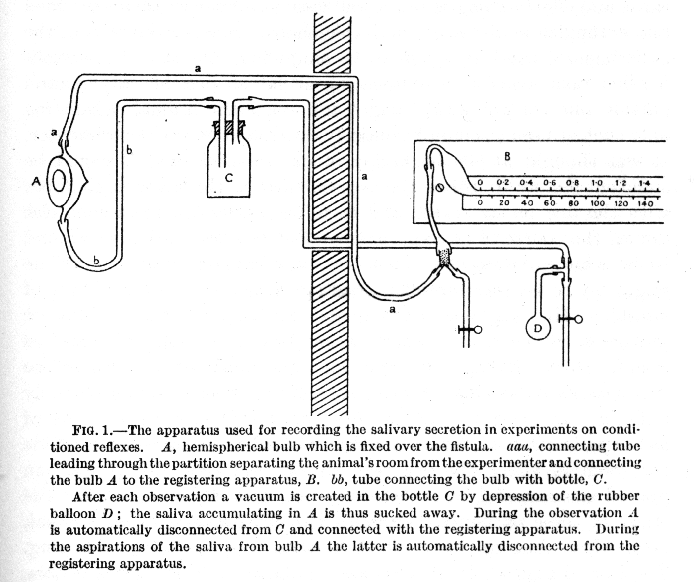
To come to the general technique of the experiments, it is important to remember that our research deals with the highly specialized activity of the cerebral cortex, a signalizing apparatus of tremendous complexity and of most exquisite sensitivity, through which the [p. 20] animal is influenced by countless stimuli from the outside world. Every one of these stimuli produces a certain effect upon the animal, and all of them taken together may clash and interfere with, or else reinforce, one another. Unless we are careful to take special precautions the success of the whole investigation may be jeopardized, and we should get hopelessly lost as soon as we began to seek for cause and effect among so many and various influences, so intertwined and entangled as to form a veritable chaos. It was evident that the experimental conditions had to be simplified, and that this simplification must consist in eliminating as far as possible any stimuli outside our control which might fall upon the animal, admitting only such stimuli as could be entirely controlled by the experimenter. It was thought at the beginning of our research that it would be sufficient simply to isolate the experimenter in the research chamber with the dog on its stand, and to refuse admission to anyone else during the course of an experiment. But this precaution was found to be wholly inadequate, since the experimenter, however still he might try to be, was himself a constant source of a large number of stimuli. His slightest movements-blinking of the eyelids or movement of the eyes, posture, respiration and so on-all acted as stimuli which, falling upon the dog, were sufficient to vitiate the experiments by making exact interpretation of the results extremely difficult. In order to exclude this undue influence on the part of the experimenter as far as possible, he had to be stationed outside the room in which the dog was placed, and even this precaution proved unsuccessful in laboratories not specially designed for the study of these particular reflexes. The environment of the animal, even when shut up by itself in a room, is perpetually changing. Footfalls of a passer-by, chance conversations in neighbouring rooms, slamming of a door or vibration from a passing van, street-cries, even shadows cast through the windows into the room, any of these casual uncontrolled stimuli falling upon the receptors of the dog set up a disturbance in the cerebral hemispheres and vitiate the experiments. To get over all these disturbing factors a special laboratory was built at the Institute of Experimental Medicine in Petrograd, the funds being provided by a keen and public-spirited Moscow business man. The primary task was the protection of the dogs from uncontrolled extraneous stimuli, and this was effected by surrounding the building with an isolating trench and employing other special structural devices. Inside the building all the research rooms [p. 21] (four to each floor) were isolated from one another by a, cross-shaped corridor ; the top and ground floors, where these rooms were situated, were separated by an intermediate floor. Each research room was carefully partitioned by the use of sound-proof materials into two compartments -- one for the animal, the other for the experimenter. For stimulating the animal, and for registering the corresponding reflex response, electrical methods or pneumatic transmission were used. By means of these arrangements it was possible to get something of that stability of environmental conditions so essential to the carrying out of a successful experiment.
Another point should be mentioned-although in this respect the means at our disposal still leave something to be desired. In analysing the exceedingly complex influence of the external environment upon the animal, the experimenter must be able to exercise full control over all the conditions obtaining during the course of any experiment. He should therefore have at his disposal various instruments for affecting the animal by different kinds of stimuli, singly or combined, so as to imitate simple natural conditions. But we were often handicapped by the conditions in which we had to work and by the shortcomings of the instruments at our disposal, for we always found that the cerebral hemispheres were sensitive to far finer gradations of stimulus than we could furnish.
It is possible that the experimental conditions I have described may raise somewhere the objection of being abnormal and artificial. However it is hardly likely, in view of the infinite variety of stimuli met with under natural conditions, that we shall hit on one that is quite unprecedented in the life of the animal. Moreover, in dealing with any phenomenon of vast complexity it is absolutely necessary to isolate the different single factors involved, so as to study them independently, or in arbitrary groups in which we can keep the individual units under control. But as a matter of fact the same objection and the same answer apply equally to the whole of animal physiology. For instance, the methods of vivisection and of the study of isolated organs and tissues, which aim at the same isolation of different individual functions, have been constantly employed, and we may safely say that the greater part of the achievements of physiology are due to the successful application of such methods of control. In our experiments it is the whole animal which is placed under a limited number of rigidly defined conditions, and [p. 22] only by this method is it possible to study the reflexes independently of one another.
The foregoing remarks give an idea of our general aim and of the technical side of our methods. I propose to introduce you to the first and most elementary principles of the subject matter of our research by means of a few demonstrations:
Demonstration. -- The dog used in the following experiment has been operated upon as described previously. It can be seen that so long as no special stimulus is applied the salivary glands remain quite inactive. But when the sounds from a beating metronome are allowed to fall upon the ear, a salivary secretion begins after 9 seconds, and in the course of 45 seconds eleven drops have been secreted. The activity of the salivary gland has thus been called into play by impulses of sound -- a stimulus quite alien to food. This activity of the salivary gland cannot be regarded as anything else than a component of the alimentary reflex. Besides the secretory, the motor component of the food reflex is also very apparent in experiments of this kind. In this very experiment the dog turns in the direction from which it has been customary to present the food and begins: to lick its lips vigorously.
This experiment is an example of a central nervous activity depending on the integrity of the hemispheres. A decerebrate dog would never have responded by salivary secretion to any stimulus of the kind. It is obvious also that the underlying principle of this activity is signalization. The sound of the metronome is the signal for food, and the animal reacts to the signal in the same way as if it were food; no distinction can be observed between the effects produced on the animal by the sounds of the beating metronome and showing it real food.
Demonstration. -- Food is shown to the animal. The salivary secretion begins after 5 seconds, and six drops are collected in the course of 15 seconds. The effect is the same as that observed with the sounds of the metronome. It is again a case of signalization, and is due to the activity of the hemispheres. That the effect of sight and smell of food is not due to an inborn reflex, but to a reflex which has been acquired in the course of the animal's own individual existence, was shown by experiments carried out by Dr. Zitovich in the laboratory of the late Prof. Vartanov. Dr. Zitovich took several young puppies away from their mother and fed them for considerable time only on milk. When the [p. 23] puppies were a few months old he established fistulae of their salivary ducts, and was thus able to measure accurately thc secretory activity of the glands. He now showed these puppies some solid food -- bread or meat -- but no secretion of saliva was evoked. It is evident, therefore, that the sight of food does not in itself act as a direct stimulus to salivary secretion. Only after the puppies have been allowed to eat bread and meat on several occasions does the sight or smell of these foodstuffs evoke the secretion.
The following experiment serves to illustrate the activity of the salivary gland as an inborn reflex in contrast to signalization:
Demonstration. -- Food is suddenly introduced into the dog's mouth; secretion begins in 1 to 2 seconds. The secretion is brought about by the physical and chemical properties of the food itself acting upon receptors in the mucous membrane of the mouth and tongue. It is purely reflex.
This comparatively simple experiment explains how a decerebrate dog can die of starvation in the midst of plenty, for it will only start eating if food chances to come into contact with its mouth or tongue. Moreover, the elementary nature of the inborn reflexes, with their limitations and inadequacy, are clearly brought out in these experiments, and we are now able to appreciate the fundamental importance of those stimuli which have the character of signals.
Our next step will be to consider the question of the nature of signalization and of its mechanism from a purely physiological point of view. It has been mentioned already that a reflex is an inevitable reaction of the organism to an external stimulus, brought about along a definite path in the nervous system. Now it is quite evident that in signalization all the properties of a reflex are present. In the first place an external stimulus is required. This was given in our first experiment by the sounds of a metronome. These sounds falling on the auditory receptor of the dog caused the propagation of an impulse along the auditory nerve. In the brain the impulse was transmitted to the secretory nerves of the salivary glands, and passed thence to the glands, exciting them to active secretion. It is true that in the experiment with the metronome an interval of several seconds elapsed between the beginning of the stimulus and the beginning of the salivary secretion, whereas the time interval for the inborn reflex secretion was only 1 to 2 seconds. The longer latent period was, however, due to some special conditions of the experiment, as will come out more clearly as we proceed. But [p. 24] generally speaking the reaction to signals under natural conditions is as speedy as are the inborn reflexes. We shall be considering the latent period of signalization in fuller detail in a further lecture.
In our general survey we characterized a reflex as a necessary reaction following upon a strictly definite stimulus under strictly defined conditions. Such a definition holds perfectly true also for signalization; the only difference is that the type of thc effective reaction to signals depends upon a greater number of conditions. But this does not make signalization differ fundamentally from the better known reflexes in any respect, since in the latter, variations in character or force, inhibition and absence of reflexes, can also be traced to some definite change in the conditions of the experiment.
Thorough investigation of the subject shows that accident plays no part whatever in the signalizing activity of the hemispheres, and all experiments proceed strictly according to plan. In the special laboratory I have described, the animal can frequently be kept under rigid experimental observation for 1 to 2 hours without a single drop of saliva being secreted independently of stimuli applied by the observer, although in the ordinary type of physiological laboratory experiments are very often distorted by the interference of extraneous and uncontrolled stimuli.
All these conditions leave no grounds for regarding the phenomena which we have termed "signalization" as being anything else than reflex. There is, however, another aspect of the question which at a first glance seems to point to an essential difference between the better known reflexes and signalization. Food, through its chemical and physical properties, evokes the salivary reflex in every dog right from birth, whereas this new type claimed as reflex -- "the signal reflex" -- is built up gradually in the course of the animal's own individual existence. But can this be considered as a fundamental point of difference, and can it hold as a valid argument against employing the term "reflex" for this new group of phenomena ? It is certainly a sufficient argument for making a definite distinction between the two types of reflex and for considering the signal reflex in a group distinct from the inborn reflex. But this does not invalidate in any way our right logically to term both "reflex," since the point of distinction does not concern the character of the response on the part of the organism, but only the mode of formation of the reflex mechanism. We may take the telephonic installation as an illustration. Communication can be effected in two ways. [p. 25] My residence may be connected directly with the laboratory by a private line, and I may call up the laboratory whenever it pleases me to do so; or on the other hand, a connection may have to be made through the central exchange. But the result in both cases is the same. The only point of distinction between the methods is that the private line provides a permanent and readily available cable, while the other line necessitates a preliminary central connection being established. In the one case the communicating wire is always complete, in the other case a small addition must be made to the wire at the central exchange. We have a similar state of affairs in reflex action. The path of the inborn reflex is already completed at birth; but the path of the signalizing reflex has still to be completed in the higher nervous centres. We are thus brought to consider the mode of formation of new reflex mechanisms. A new reflex is formed inevitably under a given set of physiological conditions, and with the greatest ease, so that there is no need to take the subjective states of the dog into consideration. With a complete understanding of all the factors involved, the new signalizing reflexes are under the absolute control of the experimenter; they proceed according to as rigid laws as do any other physiological processes, and must be regarded as being in every sense a part of the physiological activity of living beings. I have termed this new group of reflexes conditioned reflexes to distinguish them from the inborn or unconditioned reflexes. The term "conditioned" is becoming more and more generally employed, and I think its use is fully justified in that, compared with the inborn reflexes, these new reflexes actually do depend on very many conditions, both in their formation and in the maintenance of their physiological activity. Of course the terms "conditioned" and "unconditioned" could be replaced by others of arguably equal merit. Thus, for example, we might retain the term "inborn reflexes," and call the new type "acquired reflexes"; or call the former "species reflexes " since they are characteristic of the species, and the latter "individual reflexes" since they vary from animal to animal in a species, and even in the same animal at different times and under different conditions. Or again we might call the former "conduction reflexes "and the latter "connection reflexes."
There should be no theoretical objection to the hypothesis of the formation of new physiological paths and new connections within the cerebral hemispheres. Since the especial function of the central [p. 26] nervous system is to establish most complicated and delicate correspondences between the organism and its environment we may not unnaturally expect to find there, on the analogy of the methods used by the technician in everyday experience, a highly developed connector system superimposed on a conductor system. The physiologist certainly should not object to this conception seeing that he has been used to employing the German conception of "Bahnung," which means a laying down of fresh physiological paths in the centres. Conditioned reflexes are phenomena of common and widespread occurrence: their establishment is an integral function in everyday life. We recognize them in ourselves and in other people or animals under such names as "education," "habits," and "training; " and all of these are really nothing more than the results of an establishment of new nervous connections during the post-natal existence of the organism. They are, in actual fact, links connecting definite extraneous stimuli with their definite responsive reactions. I believe that the recognition and the study of the conditioned reflex will throw open the door to a true physiological investigation probably of all the highest nervous activities of the cerebral hemispheres, and the purpose of the present lectures is to give some account of what we have already accomplished in this direction.
We come now to consider the precise conditions under which new conditioned reflexes or new connections of nervous paths are established. The fundamental requisite is that any external stimulus which is to become the signal in a conditioned reflex must overlap in point of time with the action of an unconditioned stimulus. In the experiment which I chose as my example the unconditioned stimulus was food. Now if the intake of food by the animal takes place simultaneously with the action of a neutral stimulus which has been hitherto in no way related to food, the neutral stimulus readily acquires the property of eliciting the same reaction in the animal as would food itself. This was the case with the dog employed in our experiment with the metronome. On several occasions this animal had been stimulated by the sound of the metronome and immediately presented with food -- i.e. a stimulus which was neutral of itself had been superimposed upon the action of the inborn alimentary reflex. We observed that, after several repetitions of the combined stimulation, the sounds from the metronome had acquired the property of stimulating salivary secretion and of evoking the motor reactions characteristic of the alimentary reflex. The first [p.27] demonstration was nothing but an example of such a conditioned stimulus in action. Precisely the same occurs with the mild defence reflex to rejectable substances. Introduction into the dog's mouth of a little of an acid solution brings about a quite definite responsive reaction. The animal sets about getting rid of the acid, shaking its head violently, opening its mouth and making movements with its tongue. At the same time it produces a copious salivary secretion. The same reaction will infallibly be obtained from any stimulus which has previously been applied a sufficient number of times while acid was being introduced into the dog's mouth. Hence a first and most essential requisite for the formation of a new conditioned reflex lies in a coincidence in time of the action of any previously neutral stimulus with some definite unconditioned stimulus. Further, it is not enough that there should be overlapping between the two stimuli; it is also and equally necessary that the conditioned stimulus should begin to operate before the unconditioned stimulus comes into action.
If this order is reversed, the unconditioned stimulus being applied first and the neutral stimulus second, the conditioned reflex cannot be established at all. Dr. Krestovnikov performed these experiments with many different modifcations and controls, but the effect was always the same. The following are some of his results: In one case 427 applications were made in succession of thc odour of vanillin together with the introduction of acid into the dog's mouth, but the acid was always made to precede the vanillin by some 5 to 10 seconds. Vanillin failed to acquire the properties of a conditioned stimulus. However, in the succeeding experiment, in which the order of stimuli was reversed, the odour, this time of amyl acetate, became an effective conditioned stimulus after only 20 combinations. With another dog the loud buzzing of an electric bell set going 5 to 10 seconds after administration of food failed to establish a conditioned alimentary reflex even after 374 combinations, whereas the regular rotation of an object in front of the eyes of the animal, the rotation beginning before the administration of food, acquired the properties of a conditioned stimulus after only 5 combinations. The electric buzzer set going before the administration of food established a conditioned alimentary reflex after only a single combination.
Dr. Krestovnikov's experiments were carried out on five dogs, and the result was always negative when the neutral stimulus was [p. 28] applied, whether 10 seconds, 5 seconds or only a single second after the beginning of the unconditioned stimulus. During all these experiments not only the secretory reflex but also the motor reaction of the animal was carefully observed, and these observations always corroborated one another. We thus see that the first set of conditions required for the formation of a new conditioned reflex encompasses the time relation between the presentation of the unconditioned stimulus and the presentation of that agent which has to acquire the properties of a conditioned stimulus.
As regards the condition of the hemispheres themselves, an alert state of the nervous system is absolutely essential for the formation of a new conditioned reflex. If the dog is mostly drowsy during the experiments, the establishment of a conditioned reflex becomes a long and tedious process, and in extreme cases is impossible to accomplish. The hemispheres must, however, be free from any other nervous activity, and therefore in building up a new conditioned reflex it is important to avoid foreign stimuli which, falling upon the animal, would cause other reactions of their own. If this is not attended to, the establishment of a conditioned reflex is very difficult, if not impossible. Thus, for example, if the dog has been so fastened up that anything causes severe irritation, it does not matter how many times the combination of stimuli is repeated, we shall not be able to obtain a conditioned reflex. A somewhat similar case was described in the first lecture-that of the dog which exhibited the freedom reflex in an exaggerated degree. It can also be stated as a rule that the establishment of the first conditioned reflex in an animal is usually more difficult than the establishment of succeeding ones. It is obvious that this must be so, when we consider that even in the most favourable circumstances the experimental conditions themselves will be sure to provoke numerous different reflexes -- i.e. will give rise to one or other disturbing activity of the hemispheres. But this statement must be qualified by remarking that in cases where the cause of these uncontrolled reflexes is not found out, so that we are not able to get rid of them, the hemispheres themselves will help us. For if the environment of the animal during the experiment does not contain any powerful disturbing elements, then practically always the extraneous reflexes will with time gradually and spontaneously weaken in strength.
The third factor determining the facility with which new conditioned reflexes can be established is the health of the animal. A [p. 29] good state of health will ensure the normal functioning of the cerebral hemispheres, and we shall not have to bother with the effects of any internal pathological stimuli.
The fourth, and last, group of conditions has to dg with the properties of the stimulus which is to become conditioned, and also with the properties of the unconditioned stimulus which is selected. Conditioned reflexes are quite readily formed to stimuli to which the animal is more or less indifferent at the outset, though strictly speaking no stimulus within the animal's range of perception exists to which it would be absolutely indifferent. In a normal animal the slightest alteration in the environment -- even the very slightest sound or faintest odour, or the smallest change in intensity of illumination -- immediately evokes the reflex which I referred to in the first lecture as the investigatory reflex -- "What is it?" -- manifested by a very definite motor reaction. However, if these neutral stimuli keep recurring, they spontaneously and rapidly weaken in their effect upon the hemispheres, thus bringing about bit by bit the removal of this obstacle to the establishment of a conditioned reflex. But if the extraneous stimuli are strong or unusual, the formation of a conditioned reflex will be difficult, and in extreme cases impossible.
It must also be remembered that in most cases we are not acquainted with the history of the dog before it came into the laboratory, and that we do not know what sort of conditioned reflexes have been established to stimuli which appear to be of the simplest character. But in spite of this we have, in a large number of cases, found it possible to take a strong stimulus which evoked some strong unconditioned response of its own, and still succeed in converting it into a conditioned stimulus for another reflex. Let us take for example a nocuous stimulus, such as a strong electric current or wounding or cauterization of the skin. These are obviously stimuli to vigorous unconditioned defence reflexes. The organism responds by a violent motor reaction directed towards removal of the nocuous stimulus or to its own removal from it. But we may, nevertheless, make use even of these stimuli for the establishment of a new conditioned reflex. Thus in one particular experiment a strong nocuous stimulus -- an electric current of great strength -- was converted into an alimentary conditioned stimulus, so that its application to the skin did not evoke the slightest defence reaction. Instead, the animal exhibited a well-marked alimentary conditioned reflex, turning its [p. 30] head to where it usually received the food and smacking its lips, at the same time producing a profuse secretion of saliva. The following is s record taken from a research by Dr. Eroféeva:
Similar results were obtained from dogs in which cauterization or pricking of the skin deep enough to draw blood was made to acquire the properties of an alimentary conditioned stimulus. These experiments have been apt to upset very sensitive people; but we have been able to demonstrate, though without any pretension of penetrating into the subjective world of the dog, that they were labouring under a false impression. Subjected to the very closest scrutiny, not even the tiniest and most subtle objective phenomenon usually exhibited by animals under the influence of strong injurious stimuli can be observed in these dogs. No appreciable changes in the pulse or in the respiration occur in these animals, whereas such changes are always most prominent when the nocuous stimulus has not been converted into an alimentary conditioned stimulus. Such a remarkable phenomenon is the result of diverting the nervous impulse from one physiological path to another. This transference is dependent, however, upon a very definite condition -- namely, upon the relative strengths of the two unconditioned reflexes.
Successful transformation of the unconditioned stimulus for one reflex into the conditioned stimulus for another reflex can be brought about only when the former reflex is physiologically weaker and biologically of less importance than the latter. We are led to this conclusion from s consideration of Dr. Eroféeva's experiments. [p. 31] A nocuous stimulus applied to the dog's skin was transformed into a conditioned stimulus for the alimentary reflex. This, we consider, was due to the fact that the alimentary reflex is in such cases stronger than the defence reflex. In the same way we all know that when dogs join in a scuffle for food they frequently sustain skin wounds, which however play no dominant part as stimuli to any defence reflex, being entirely subordinated to the reflex for food. Nevertheless there is a certain limit -- there are stronger reflexes than the alimentary reflex. One is the reflex of self-preservation, of existence or non-existence, life or death. To give only one example, it was found impossible to transform a defence reaction into an alimentary conditioned reflex when the stimulus to the unconditioned defence reaction was a strong electric current applied to skin overlying bone with no muscular layer intervening. This signifies that the afferent nervous impulses set up by injury to the bone, and signalizing far greater danger than those set up by injury to the skin, cannot acquire even a temporary connection with the part of the brain from which the alimentary reflex is controlled. Nevertheless, an the whole, the foregoing considerations emphasize the advantage of using the alimentary reflex for most of our experiments, since in the hierarchy of reflexes this holds a very high place.
While, as we have seen, very strong and even specialized stimuli can under certain conditions acquire the properties of conditioned stimuli, there is, on the other hand, a minimum strength below which stimuli cannot be given conditioned properties. Thus a thermal stimulus of 45°C. applied to the skin can be made into an alimentary conditioned reflex, whereas at 38° to 39°C. (approximately 2°C. above the skin temperature in the dog) a thermal stimulus is ineffective [experiments of Dr. Solomonov]. Similarly, while with the help of a very strong unconditioned stimulus it is possible to convert a very unsuitable stimulus -- for example, one which naturally evokes a different unconditioned reflex -- into a conditioned stimulus, it is exceedingly difficult or even impossible with the help of only a weak unconditioned stimulus to transform even a very favourable neutral stimulus into a conditioned stimulus. Even where such a conditioned reflex is successfully established, its occurrence results only in a very small reflex response. Some unconditioned stimuli may be permanently weak, others may display a weakness which is only temporary -- varying with the condition of the animal. As an example of the last we may take food. In the hungry animal food [p. 32] naturally brings about a powerful unconditioned reflex, and the conditioned reflex develops quickly. But in a dog which has not long been fed the unconditioned stimulus has only a small effect, and alimentary conditioned reflexes either are not formed at all or are established very slowly.
By complying with all the conditions which I have enumerated -- which is not a very difficult task -- a new conditioned reflex is infallibly obtained. We apply to the receptors of the animal rigidly defined stimuli; these stimuli necessarily result in the formation of a new connection in the hemispheres with a consequent development of a typical reflex reaction.
To sum up, we may legitimately claim the study of the formation and properties of conditioned reflexes as a special department of physiology. There is no reason for thinking about all these events in any other way, and it is my belief that in these questions prejudices blunt the intellect and that generally speaking the preconceptions of the human mind stand in the way of any admission that the highest physiological activity of the hemispheres is rigidly determined. The difficulty is mainly due to the tremendous complexity of our subjective states; and, of course, these cannot yet be traced to their primary causations.


Footnotes
[1] Mendeléef's cement: Colophonium, 50 grammes; ferric oxide, 40 grammes; yellow beeswax, 25 grammes.
[2] In almost all the experiments quoted in these lectures the amount of salivary secretion is, for the sake of uniformity, given in drops. It was, however, only in the very earliest period of the research -- before the separation of the experimenter from the animal was made -- that the actual number of drops falling from a small funnel fixed over the fistula was counted, and only a few of these experiments are given. In the great majority of the experiments the salivary secretion was measured by the displacement of water in a graduated tube or by the electric recorder, allowing much greater accuracy of measurement. The readings so obtained have been converted, in the tables, into drops. Thus, in some experiments it will be noticed that the number of drops is given to an accuracy of one-tenth.