
(Return to index)
Posted July 2001
[Classics Editor's note: Pavlov
used both square and round brackets in his texts. These have been preserved
but can lead to confusions as to which insertions are the author's and
which are the editor's. Page numbers, reference numbers, and the occasional
"sic" have been inserted in square brackets by the Classics editor.
All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE XXII
The general characteristics of the present investigation and its
special difficulties
-- Discovery of certain errors necessitating the modification of
some earlier interpretations.
A scientific investigation of biological phenomena can be conducted along several different lines each of which would treat the problem from a different point of view. For instance, one may have in view the purely physico-chemical aspect, analysing the elements of life by the methods of physics and chemistry. Again, keeping in view the fact of evolution of living matter one can try to elucidate the functions of complex biological structures by studying the functions of individual cells and of elementary organisms. Finally, one can make an attempt to elucidate the activities of complex structures in their fullest range directly, seeking for rigid laws governing this activity, or, in other words, trying to define all those conditions which determine the form this activity takes at every instant and in all its variations. The line of inquiry which has been adopted in the present investigation obviously belongs to the third point of view. In this research we were not concerned with the ultimate nature of excitation and inhibition as such. We took them as two fundamental properties, the two most important manifestations of activity, of the living nervous elements. Nor was it our aim to interpret the activity of the hemispheres in terms of elementary functions of the nervous system, as has been done, for example, in the physiology of the nerve fibres. We intentionally neglected also the controversial problem of the actual localization of these two fundamental processes, and did not attempt to assign them to either of the two elements of the nervous structure, namely, the nerve cell and the synaptic junction or fibrillary connection between two individual nerve cells. The acceptance of the more general conception of the two processes of inhibition and excitation as the basic functions of the nervous cellular structures was sufficient for the purposes of our research, the study of conditioned reflexes being of the nature [p. 378] of a general investigation of the functions of the cellular structures of the cortex as exhibited in various reactions of the organism to a multitude of separate stimuli which originate from within or from without the organism -- stimuli for the reception of which there is such an unbounded number of separate cortical cells and which after extirpation of the cortex lose their significance for the organism. It is highly probable that excitation and inhibition, the two functions of the nerve cell which are so intimately interwoven and which so constantly supersede each other, may, fundamentally, represent only different phases of one and the same physico-chemical process. The primary aim of our research was the accurate determination and tabulation of different phases of the cortical activity -- the absence or presence of an inhibitory or excitatory phase, the exact conditions under which the intensity of the excitatory or inhibitory process varied, and the mutual interrelation between these processes. It is obvious that in its intrinsic nature our work is closely allied to the work of Sherrington and his co-workers upon the spinal cord, and it is impossible not to notice in how many points the different aspects of the nervous activity of the cortex correspond with those described in the physiology of the spinal cord; a fact which seems strong evidence of a similarity of the fundamental laws governing the nervous activity in the two cases.
Research upon the activity of the cortex along these lines must unavoidably present exceptional difficulties. The extraordinary reactivity of the cortex, on the one hand, and the unbounded volume of stimuli continually pouring into it, on the other hand, are responsible for the two fundamental peculiarities of the cortical activity -- namely, first, that it is determined in every minutest detail, and second, that it is in a state of perpetual flux, changing so rapidly that it becomes practically impossible to observe any aspect of it in an entirely pure and uncontaminated form and to appraise and control all the determining conditions. The minutest changes in the environment or inside the organism itself -- changes which may be imperceptible to us and unsuspected -- have a profound effect upon the cortical activities. It is obvious that these special peculiarities of the research are in many instances the cause of fallacies, especially since it is so tempting to adhere to different fancied analogies and plausible generalizations -- a tendency which cannot be too much guarded against in the present state of the research. The mind, so to speak, often fails to keep pace with the tremendous variety of [p. 379] interrelations, and this is why our interpretations were often too limited and led to errors which have had to be constantly corrected. Indeed I have no doubt that the presentation of the subject-matter attempted in these lectures will in the future still be corrected in many details. Errors in interpretation, and errors sometimes in the methods of observation, are naturally to be expected in a study of such astounding complexity. These special peculiarities of our subject is the reason why I thought it advisable to delay a systematic presentation of our prolonged researches until now: new problems are perpetually arising, and at the same time an equally large number of questions are still left unsettled. We often feel compelled to turn our attention from problems which directly confront us to some unexpected new phenomena which introduce fresh problems or which necessitate a revision of old points of view. This general aspect of the investigation into the cortical activity I want particularly to emphasize in the present lecture, taking for my examples some fresh observations which have not been discussed in the preceding lectures.
The surprising minuteness of detail in which the cortical activity is determined by external and internal agencies, and the extraordinary precision and delicacy of the responsiveness of the cortex to even the slightest changes in these agencies, are clearly illustrated by the following observations, which are both taken from the later period of our work.
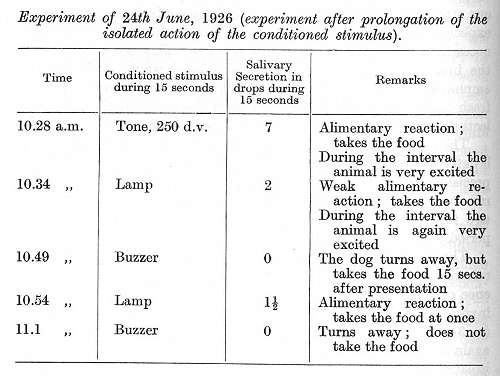
The dog used in the first of these experiments has already been referred to in the preceding lectures. It passed through the experience of the great flood in Petrograd, and afterwards served for an investigation of a functional disturbance of the acoustic analyser (p. 316). In the course of a month in which the animal behaved again normally a differentiation of pitch was developed. During the isolated action of the conditioned stimulus (10 seconds) the secretory effect was as much as 5 drops; the difference between the effects of strong and weak stimuli was very definite, and the food given in reinforcement was always taken with avidity. In the stand the animal stood quietly. We now introduced an apparently very small modification in the experiment, the isolated action of the conditioned stimulus being prolonged a further 5 seconds. As a result of this the entire conditioned activity was immediately disturbed. The following are illustrative experiments taken from the two periods of the research: [p. 380]
The first application of the conditioned stimulus in the experiment of 24th June produced a greater secretory effect than usual, for the obvious reason that the isolated action of the conditioned stimulus had been protracted for a further 5 seconds. The alimentary motor reaction in response to the stimulus was lively, the food being consumed at once; everything promised a normal experiment, and there were so far no indications of any special deviation from normal. However, already during the interval between the first and second stimuli the animal showed an unusual state of excitation. This was [p. 381] followed later on by an obvious paradoxical phase : a strong stimulus (buzzer) failed to elicit the secretory reaction while the motor reflex considerably diminished. The animal on the first application of the buzzer (10.49 a.m.) accepted the food after some delay, and on the second application (11.1 a.m.) did not touch the food at all. A weak stimulus (lamp) nevertheless continued to evoke a secretion (although a diminished one) and to evoke a motor alimentary response. We returned to the usual length of isolated action of the conditioned stimulus (10 seconds) on the following day. The disturbance, however, became still more pronounced, the secretory effect being absent throughout the experiment, while the animal turned to the food only after weak stimuli and would not touch it after strong ones. On the third day all reflexes returned to normal, except that the tone which had been the first stimulus employed on the day (24th June) when the isolated action of the stimuli was prolonged, still gave a diminished secretory effect (only a half of its usual secretion during 10 seconds). A further experiment with a temporary prolongation of the isolated action of the conditioned stimuli produced exactly the same pathological state (paradoxical phase) of the animal. These observations provide a brilliant example of the exquisite delicacy in the reactivity of the cortex, showing how considerable may be the effect of such minute changes in the conditions of an experiment.
The second dog, also previously mentioned, -- "Brains" -- was of an extremely inhibitable type, and the same change in the experimental conditions led to exactly opposite results. This dog when left in the stand without application of any stimuli quickly became drowsy, with the result that not only conditioned reflexes disappeared but also reflexes in response to the actual administration of food. For the purpose of overcoming this drowsiness we had used the usual method of abbreviating the isolated action of the conditioned stimulus to 1/2-1 second. After three weeks of this practice the drowsiness disappeared, and the dog now took the food immediately on presentation and consumed it with avidity. A prolongation of the isolated action of the conditioned stimulus to 5 seconds revealed the presence of a conditioned secretory reflex. On continuation of the experiments with the isolated action of 5 seconds the reflex maintained its strength for several days. After this the reflex again diminished and the animal once more succumbed to drowsiness. However, it was now sufficient to prolong the conditioned stimulus [p. 382] to 10 seconds for the animal to become again alert; the secretory effect returned after several repetitions of the prolonged stimulus and, what is important, it appeared within the first 5 seconds of the isolated action of the conditioned stimulus. On continuation of the experiments with the isolated action of 10 seconds the animal again became drowsy and the secretory conditioned reflex again disappeared. A further prolongation of the conditioned stimulus to 15 seconds produced a similar effect to that of the first prolongation to 10 seconds, i.e. a temporary return of the alert state and a considerable conditioned secretion starting well within the first 5 seconds. The same sequence of events was repeated yet twice more on prolongation of the conditioned stimulus to 20 and 25 seconds. The following is an example of an experiment:

On the next day the isolated action of the conditioned stimuli was prolonged to 20 seconds. The following is an experiment after two days' practice with the longer interval:

Thus every small prolongation of the isolated action of the conditioned stimulus evoked after a few repetitions a temporary state of excitation of the dog: the drowsiness disappeared, and the secretory reaction reappeared, starting soon after the beginning of the conditioned stimulus and well within the period in which there was no secretion during the shorter isolated action of the stimulus. [p. 383]
The latter fact shows that the enhanced secretion was not merely due to the prolongation of the period of observation but was the genuine result of a cortical excitation.
We thus see that one and the same change led to opposite effects in the two dogs: in the first it resulted in inhibition, in the second it abolished inhibition. Most probably the difference in the effects was dependent on the fact that the first animal was alert during the experiments (i.e. in a state of cortical excitation), while the other dog was drowsy (i.e. in a state of cortical inhibition).
We are constantly confronted by cases in which pathological changes in the state of the animal are brought about by some unknown secondary conditions. It has repeatedly been mentioned in the preceding lectures that conditioned reflexes to stimuli belonging to different analysers are normally of different magnitude. Remembering that the nervous process is considered by present-day physiologists to be identical in all nerve fibres, and having somehow conceived the idea that the difference in our perception of light, sound, etc., must have some physical basis in differences of corresponding cortical elements, we were for a considerable time inclined to attribute the difference in the magnitude of the conditioned effect to individual peculiarities of the cells of the different analysers. Reinvestigation of the whole question, as has been mentioned already, showed that the differences in the magnitude of the conditioned reactions to stimuli belonging to different analysers, depend fundamentally upon the intensity of the stimuli themselves (p. 269). This conclusion, it will be remembered, was arrived at in the following manner. We knew for a long time that in a conditioned compound stimulus made up of two agencies belonging to different analysers one of the stimuli almost invariably overshadows the other, as can easily be revealed by testing the individual components separately. In such a compound our usual auditory stimuli, in the majority of dogs, over-shadowed visual, tactile and thermal components, but in a compound made up of a weakened auditory stimulus and a strong visual one the relations were entirely reversed. This observation proved the difference in response to be due not to fundamental differences in the cellular structure of the various analysers, but to the relative intensity of the individual stimuli. Now, although the above holds good in the majority of cases, in some few animals the difference in the magnitude of the conditioned response to different agencies was found to be entirely absent. We have already directed considerable [p. 384] attention to the study of this deviation, but we are not yet in a position to state definitely under what special conditions it occurs. Some of the determining conditions are, however, known. These special cases depend to some extent upon the general type of the nervous system of the animal -- excitable or inhibitable. In the inhibitable type the usual relation between intensity of stimulus and magnitude of effect is especially obvious, occurring practically without exception otherwise than in extreme pathological states. It is true that if the period of isolated action is short there may not be any apparent difference between the effects of strong and weak stimuli, since in the initial stage the effect is practically the same in both cases; but with a prolongation of the stimulus the secretion augments rapidly in the case of strong stimuli and only slowly in the case of weak stimuli. In the case of excitable and very greedy dogs it is, on the contrary, the abbreviation of the conditioned stimulus which helps to disclose the usual relation between the magnitudes of the effects of weak and of strong stimuli, a relation which may not be apparent when the stimuli are more prolonged.
On account of the immense number of different conditions determining the different states of activity of the cortex I feel that even now the physiological analysis of many seemingly simple and well-known facts is very often far from perfect. I will describe, for example, the observations upon one of our recently acquired animals [experiments of Drs. Podkopaev and Virjikovsky]. In this case the conditioned reflexes to different agencies were developed with the following important variation in the usual method. The first agent to which conditioned properties were to be given was applied alternately with and without reinforcement by food. The conditioned reflex developed comparatively quickly (by the 20th application). In the case of the next stimulus the reinforcement was given at every third application. The reflex developed even quicker than before (by the 7th application). The animal became, however, extremely excited. Finally a third agent was reinforced only at every fourth application, and in this case the conditioned reflex failed to develop and the animal became somewhat drowsy. The last stimulus was applied a total of 240 times (60 times in conjunction with food).
Let us, having recourse to all our previous knowledge, endeavour to interpret these facts. Why, in the last case, did the conditioned reflex fail to develop? or, at any rate, why was it for so long delayed, if, indeed, it would have developed at all? The fundamental [p. 385] mechanism of development of a conditioned reflex depends upon excitation of some definite point in the cortex coincidently with a more intense excitation of some other point, probably also of the cortex, which leads to a connection being formed between these two points; and reversely, if such a coincident stimulation of these points is not repeated for a long time the path becomes obliterated and the connection disrupted. But when once such a path has been firmly established it remains intact without further practice, for months and years. It is obvious that under suitable conditions a new connection must be formed at the very first occurrence of the simultaneous excitation and become strengthened by every repetition. In ordinary experiments with normal dogs about twenty reinforced repetitions are required to establish the first of any conditioned reflexes experimentally produced, and this number sufficed in the first variation (alternate reinforcement) described above. In the establishment of subsequent reflexes under normal conditions only three to five repetitions are necessary, and yet in the case now under discussion (reinforcement of every fourth application) sixty repetitions produced no result. The first possible explanation is the prolongation of the interval between the separate reinforcements, but this cannot be the reason in our case since with the same intervals of time, but omitting the non-reinforced applications during the intervals, the conditioned reflex invariably and quickly develops. It is obvious, therefore, that the frequent repetition of the non-reinforced agent must oppose a powerful resistance to the development of a conditioned reflex. We know already that every new stimulus which evokes the investigatory reflex ceases on repetition to have any effect unless the stimulus has been followed up with some other reflex. Such a disappearance of the effect is known to be due to a development of inhibition in those cortical elements upon which the stimulus acts. Consequently, in the third variation of the experiment such an inhibition might have developed on account of the application of three non-reinforced stimuli, the cortical elements never acquiring with the fourth, reinforced stimulus a sufficient state of excitation for a connection to be formed with the excited alimentary nervous elements. But although such an interpretation seemed highly plausible it failed when subjected to the following experimental test. When the stimulus had been repeated 240 times (80 times with reinforcement) a pre-established conditioned stimulus was applied 30 seconds after the ineffective [p. 386] stimulus. No inhibitory after-effect was ever observed. Obviously, therefore, the ineffective agent could not have produced any widespread inhibition. It was still possible, however, that on account of prolonged practice the inhibitory effect had become extremely concentrated in a narrow region; but neither was this explanation supported by our experiments, for when we now began to reinforce the agent at each successive application the conditioned secretory effect was already considerable at the third reinforcement, showing that the rate of development of the new conditioned reflex was maximal. We cannot, therefore, regard the stimulus as having had any definite inhibitory properties. Several further possible explanations have also been tested experimentally, but we have hitherto failed to disclose the nature of this phenomenon. So far, our experiments simply show that we have not yet gained command over all the renditions which determine the development of conditioned reflexes. The conditions enumerated in the second lecture, however sufficient they may be as regards the development of all those reflexes with which we have been accustomed to deal, nevertheless do not finally exhaust the subject, since having had all of them in our mind we have still failed to understand the results of the experiments described above. There must exist some further condition which has up to the present been overlooked. This failure only serves to demonstrate once more the surprising extent to which, in every detail, the cortical activity is determined, and the astonishing reactivity of the cortical elements.
It was, of course, obvious from the start that there was no immediate possibility of attacking the complex activity of the cortex from its physico-chemical aspect. Neither did there appear to be any real hope of approaching an understanding of the cortical activity through the study of the elementary properties of the nervous tissue. We have come now to see that we do not possess even yet a full descriptive knowledge of the various aspects of this activity. Our chief task in studying the cortical activity at the present time must therefore consist in reducing the tremendous mass of various separate observations to terms of a progressively diminishing number of general and more fundamental units. This we have fully realized, and in some cases we seem to be approaching our goal, but in other cases we find ourselves confronting some entirely new aspects of the cortical activity, which sometimes are within, and at other times beyond the range, of our present powers of analysis [p. 387].
In the beginning of the present research, on the basis of very definite but purely external signs, we distinguished three types of inhibition -- external inhibition, internal inhibition, and sleep. Accumulation of further observations permitted us to fuse the last two types by showing that the apparent differences are only secondary. Sleep and the various forms of internal inhibition are aspects of one and the same process, which in the one set of cases is fragmented and localized, in the other case diffused. When in the course of our further investigation of conditioned reflexes we met with the phenomenon of mutual induction, it was natural that we should perceive the similarity between negative induction and external inhibition. Hence a fundamental identity of all the three types of inhibition appeared very probable, and special efforts were directed to the collection of further evidence on the point. Some of this evidence has been given before, and fresh evidence will be added in the present lecture. In the lecture upon induction an experiment was described in which a conditioned defence reflex to a tactile stimulation of a place on the skin exerted an inhibitory after-effect upon a conditioned alimentary reflex to tactile stimulation of other places. We found reason to believe that this inhibition was in part cortical (p. 202). In the lecture upon different transition phases between the alert state and sleep it was shown that some of these phases can be observed not only during the after-effect of internal (differential) inhibition, but also under the influence of external inhibition (p. 276). I am now in a position to add some further considerations in favour of the identity of internal and external inhibition. We are inclined to regard the frequently mentioned fact of overshadowing in a compound conditioned stimulus of the weaker by the stronger component as based upon external inhibition. The cortical elements belonging to the strong stimulus inhibit those of the week stimulus, and the latter, therefore, can establish only a weak connection with the unconditioned centre. This theory is substantiated by the fact that the strengths of newly established conditioned connections depend on the relative strengths of the conditioned stimuli. This consideration obviously approximates still more the phenomenon of external inhibition to negative induction, in which the application of the positive conditioned stimulus reinforces, or even re-establishes, the inhibitory state of the nervous elements acted upon by the inhibitory stimulus. Many of my collaborators [Drs. Mishtovt, Krjishkovsky Leporsky] noticed that for a quick and complete establishment [p. 388] of a conditioned inhibitor it is of considerable importance to choose a stimulus which is not much weaker in strength than the positive component of the combination. Recent experiments performed in this connection by Dr. Poursikov not only corroborate these observations, but bring out the important fact that the external inhibition which on account of the investigatory reflex is produced at the first application of the new stimulus, often gradually and almost imperceptibly undergoes transition into the permanent conditioned inhibition. In spite of the cumulative effect of the foregoing evidence it is by no means conclusive, and I feel entitled to advance the view, that external and internal inhibition are fundamentally the same, only as a strong probability.
It was noticed in the nineteenth lecture that three different types of external stimuli lead to an inhibitory state of the cortex, namely, very weak stimuli, very strong stimuli, and unusual stimuli, and an attempt was made to give a general biological interpretation of this effect. The physiological mechanism of the effect of these stimuli is as yet obscure, neither do I find it possible to discuss the problem of inhibition in its entire range. The experimental material, although considerable, is not yet sufficient to establish any general and definite conception of the nature of inhibition and its relation to excitation. Explanations which seem to fit some one group of phenomena fail when applied to other groups. Many observations do not fit in with any of the theories, and our conception of the mechanism involved has had to be changed many times in the course of our research, never entirely satisfactorily. Here again, as in the whole of our research, we can only collect and systematize facts. The fact that very powerful and very weak stimuli have a stronger tendency than medium stimuli to produce inhibition belongs to the category of unsolved problems along with the mechanism of dis-inhibition, the positive effect of negative stimuli in the "ultra-paradoxical phase" and the negative effect of positive stimuli in the case of damage to the cortex. In many instances we fail even to see which of the phenomena are closely related to each other and which are isolated and radically different. To illustrate the difficulty of such cases I shall attempt to deal with the question of the production of inhibition by new events or a rearrangement in the grouping of old events. We change, for example, the manner in which the conditioned stimulus is reinforced by food. Instead of a plate automatically moving with food from behind a screen, [p. 389] a measured amount of food is now delivered mechanically into a stationary plate. Many dogs after this change persistently decline the food, and all conditioned reflexes disappear. This, of course, is a case of inhibition, but what explanation can be suggested as to its mechanism? It may be compared with an observation discussed in the thirteenth lecture where a change in the sequence of a series of conditioned stimuli led in some animals to a more or less profound inhibition of the entire conditioned activity -- an inhibition which lasted for several days, notwithstanding that a return was made to the previous order of stimulation. What happens here with a small number of conditioned stimuli can well be imagined to take place in respect to the entire environment. Constant repetitions of external events in a stereotyped order may lead to a definite stereotyped pattern of activity in the cortex, each new distribution of the stimuli now producing a disturbance in the pattern of the cortical activity, leading to inhibition in exactly the same manner as in our experiments with alteration in the sequence of the conditioned stimuli. This comparison does not, however, explain the mechanism of the development of inhibition in this particular case. Was it a result of the investigatory reflex due to the change in environment, or are the investigatory reflex and this very prolonged inhibition two independent phenomena? In favour of the first supposition the fact can be advanced that in very inhibitable dogs the investigatory reflex exercises an extraordinarily prolonged inhibitory after-effect.
The whole subject of conditioned reflexes has, of course, been continuously growing and expanding during the twenty-five years of its existence. The present lecture should show to the reader the exceptional difficulties we still meet with. Many similar difficulties, though some of them now seem trivial, confronted us throughout; even in our comparatively early material we still find scope for revision, and for the correction of different, sometimes important, errors. I shall discuss one of these which was discovered and corrected in repeating some old experiments during the preparation these lectures, and another which is still beng [sic] investigated.
In the fourth lecture three different modes of re-establishment extinguished conditioned reflexes were described. In the first, spontaneous recovery occurred after a longer or shorter interval of rest; such recovery was slow in developing, but stable. In the second, quick recovery was brought about with the help of reinforce- [p. 390] ment by the underlying unconditioned reflex. In the third, quick recovery was effected through the introduction of some extraneous reflex. The recoveries in the last two cases were described as fundamentally differing from one another, since the former was not only rapid, but also stable, whereas the latter, though equally quick, was only temporary -- vanishing with the disappearance of the extraneous reflex and its after-effect, when the inhibition would again acquire its full intensity and maintain itself until the final recovery occurred spontaneously, just as if no extraneous stimulus had over intervened. The term "dis-inhibition" was therefore applied only to the latter case. The difficulty of interpreting the mechanism of this difference was immediately recognized. Recently during experiments upon some new aspects of the relation between the conditioned and the unconditioned stimulus these old observations were repeated, and found to be inaccurate [experiments of Dr. Podkopaev].
The recovery of an extinguished conditioned reflex is found in both cases to be temporary -- in the case of the special reinforcing agent as well as under the influence of an extraneous reflex. In both cases the extinguished conditioned stimulus recovers its positive effect only for a time, then the positive effect again disappears, recovery occurring spontaneously. When the extinguished conditioned reflex is alimentary while the extraneous one is a defence reflex based on acid, the extent and duration of the disinhibition can be seen with great ease and regularity to be identical whichever method of restoration is employed. These experiments were conducted on two dogs, and they entirely corroborated one another. I shall describe the experiments on one of the dogs in detail. On extinguishing the conditioned alimentary reflex to a metronome it was found that the reflex remained at zero for 20 minutes counting from the last non-reinforced application of the conditioned stimulus. Then spontaneously the reflex began slowly to recover, reaching at the thirtieth minute 40% of its original value. After a fresh extinction to the first zero the conditioned stimulus was immediately reinforced, but it was again found twenty minutes later to give a zero effect. When, however, in another experiment it was tested 10 minutes after a similar reinforcement a positive effect was exhibited. Exactly similar results were obtained when acid instead of food was administered at the first zero, the reflex being again tested at the previously mentioned intervals of time. The following [p. 391] are the actual figures of some of the experiments: The conditioned stimulus on its first application on a particular day gave 6 drops in twenty seconds; the stimulus was reinforced immediately after complete extinction; tested after 10 minutes it gave 3 drops in twenty seconds. On the following day the experiment was repeated under precisely similar conditions. The stimulus gave, to start with, 7 drops during 20 seconds, and tested twenty minutes after reinforcement at the first zero of extinction gave no trace of any secretory effect. The experiments were now performed with administration of acid after the extinction of the alimentary conditioned reflex to its first zero. In the first experiment the reflex measured at the start 5 drops in twenty seconds. Tested 10 minutes after administration of acid following the first zero it gave 2 drops. On the following day the conditioned stimulus again gave 5 drops at first, but tested twenty minutes after administration of acid following the first zero it remained without any secretory effect. The maximum of the dis-inhibiting effect in both cases was reached, of course, much earlier than ten minutes from the administration of food or acid following the first zero. The error in our older experiments was due to an obvious fallacy. The comparison of the rate of recovery in the two cases had been made between the effect of the special reinforcing agent and the effect of the much weaker extraneous reflexes evoked by different auditory, visual, tactile and other stimuli which generally have only a short after-effect, while the comparison should have been made with the dis-inhibiting effect of other extraneous agencies, e.g. chemical ones, which have as long an after-effect as has food. The error was facilitated by the fallacious conception that the unconditioned stimulus underlying the conditioned one must stand in some special relation to the latter, conferring special powers of re-establishment after extinction. The result of these recent experiments inclines us more and more to believe that the inhibitory process arises in the nerve cells themselves and not in the connecting path between those cells excited by the conditioned stimulus and those excited by the special unconditioned stimulus employed. Otherwise it is difficult to reconcile the fact of the identical restorative action of the acid and food.
The second probable error I wish to describe is still undergoing investigation, but I permit myself to discuss it now, on the one hand on account of the extreme importance of the point involved, and on the other hand because it illustrates once more the exceptional [p. 392] difficulties presented in this research to the establishment of exact facts. It will be remembered that in the second lecture we discussed the essentials necessary for the establishment of conditioned reflexes. After establishing that the action of the originally neutral but potentially conditioned stimulus must overlap that of the unconditioned stimulus, we insisted also that the former must precede, by however short a time, the commencement of the latter. When the unconditioned stimulus was applied 5-10 seconds before the neutral stimulus it was found impossible to develop a conditioned reflex even by 300-400 repetitions, whereas by the usual method any conditioned reflex can be established in the average dog by so few as 3-20 repetitions. It was natural to suppose that the strong unconditioned stimulus acting on some part of the cortex evoked in virtue of external inhibition such a profound inhibitory state in the rest of the cortex that all stimuli reaching these parts became ineffective. Such a state may be compared with that of a man preoccupied with some definite activity, who remains "deaf" and "blind" to anything occurring round about him -- a familiar psychical phenomenon which is accepted from a physiological point of view as undoubtedly corresponding with an objective reality. The plausibility of the above reasoning made us confident of its validity until recently, when our point of view changed. The question was raised as to the mechanism by means of which an early reinforcement of the conditioned stimulus, i.e. a shortening of its isolated action, obstructs the development of inhibition in the cortical cells acted upon by the conditioned stimulus. In investigating this problem with a modification of the experiments we came unexpectedly on a new fact, viz. that if the unconditioned stimulus is administered before the pre-established conditioned stimulus the conditioned reflex becomes inhibited. Our attention naturally became directed to the exactly comparable case where, instead of a pre-established conditioned stimulus, we deal with a neutral agent which is intended for the development of a conditioned reflex. The effect of the unconditioned stimulus on both is precisely the same, since it exhibits in either case the properties of an external inhibitor. In contrast with these observations an introduction of a small modification in these time relations between the unconditioned stimulus and either of the other two causes the hitherto neutral agent to acquire conditioned properties and the conditioned stimulus to be strengthened in its pre-established ones. This contrast reminds [p. 393] us of another set of facts -- the relation between the development of a secondary conditioned reflex and the development of conditioned inhibition, the case in which under identical external conditions, but with a small change in the time relations between the two stimuli, there develops in the one case a process of excitation, in the other case a process of inhibition. All these observations concerning the action of the unconditioned stimulus point to the view (which involves a considerable modification of our original conception), that the mechanism of development of a conditioned reflex and the mechanism of external inhibition are somehow similar, and that the process of external inhibition bears some relation to the development of new connections between different cortical elements. If the analogy between external inhibition and the development of conditioned reflexes holds good, it should be expected that in the case where the unconditioned stimulus slightly precedes the action of the neutral agent there would in the very beginning be an opportunity for formation of a link between their respective cortical points, leading to the formation of an unstable conditioned reflex. This reflex would, however, rapidly undergo inhibition on repetition of the superimposed stimulation. The first preliminary experiments have fully confirmed our supposition. We had already noticed that administration of the unconditioned stimulus immediately preceding the pre-established conditioned one led only gradually to a definite diminution of the reflex, a diminution which was the more rapid and the more profound the smaller the intensity of the conditioned stimulus. Remembering this, we applied the similar combination of the unconditioned stimulus with the hitherto neutral stimulus, but repeated this combination only a very few times, to avoid the development of the inhibitory process. In many cases the expected result was obtained. The hitherto neutral stimulus when now tested alone revealed undoubted conditioned properties [experiments of Mlle. Pavlova and Drs. Kreps, Podkopaev, Prorokov and Koupalov]. Considering now after these preliminary experiments the results obtained in the much earlier experiments by Dr. Krestovnikov (p. 27), we found that the neutral stimuli were tested by him for a conditioned effect only after a very large number of repetitions of the unconditioned stimulus slightly preceding the neutral agent; moreover, even under these circumstances the stronger neutral stimuli when tested singly had at the first test some secretory effect. This secretory effect, however, was explained as a casual and not [p. 394] a true conditioned effect -- a brilliant illustration of the danger of too hasty generalizations. We imagined that if it were a true conditioned reflex it would increase in intensity on repetition of the combination, and not diminish end finally vanish, as happens in these experiments. It is possible also that we were misled by the absence of a definite conditioned motor reaction.
As before remarked, the problem here dealt with is being worked out in all its implications, under the strictest control and with the help of the knowledge which has been gained in the last few years. If the preliminary experiments described above should be fully upheld, an important fact in the physiology of the cortex will be disclosed -- namely, that new connections can be established in the cortex, not only in the areas of optimal excitability, but also in those areas which are in one or another phase of inhibition.
In the present lecture it has been my aim not so much to dwell on the details of the different experiments as to lay the strongest emphasis upon the fundamental peculiarities of the method of conditioned reflexes. I believe that the wealth of facts discussed in all the preceding lectures is in itself a sufficient indication that the whole problem is worthy of an intense scientific research, which should result in accumulation of a great number of valuable data. I have not, therefore, hesitated to expose in the present lecture some of the weaknesses in our own scientific venture. Full realization of the difficulties seems to me preferable to disregarding them. Moreover, it has been my desire to forewarn future workers in this field of the extraordinary complexity and the difficulties which they are bound to encounter.
On the whole, looking back upon this new field of physiological research I find it full of fascination, especially since it satisfies two of the fundamental cravings of the human intellect -- striving to realize ever new and new truths, and to protest against the pretension of finality in truth we have already gained. In this domain there will for long remain an immense breadth of uncharted ocean compared with the small patches of the known.