
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE IV
Internal inhibition of conditioned reflexes: (a) Extinction.
Towards the end of the last lecture I discussed external inhibition of conditioned reflexes, as exhibited in numerous cases of temporary clashing between conditioned reflexes and other extra excitatory processes in the brain, and we saw how this clashing led to weakening, :e or less profound, and sometimes even to the disappearance of the conditioned reflexes.
In the second type of inhibition, which may be termed internal inhibition, the positive conditioned stimulus itself becomes, under definite conditions, negative or inhibitory; it now evokes in the cells of the cortex a process of inhibition instead of the usual excitation. Conditions favouring the development of conditioned reflexes of the negative or inhibitory type are of frequent occurrence, and these reflexes are met with not less frequently than reflexes of the positive or excitatory type.
The most striking difference between external and internal inhibition is that, whereas under the conditions described in the preceding lecture external inhibition is produced on the very first application of an extra stimulus, internal inhibition on the other hand always develops progressively, quite often very slowly, and in many cases with difficulty.
I shall start by describing that form of internal inhibition which was first encountered in our researches, and shall trace the growth of our present conception of its nature.
Demonstration.-- The following is an example of an experiment illustrating the first group of internal inhibitions.
We are taking for this experiment the same dog that was used in the second lecture for the conditioned reflex to the sound of the metronome. In testing the reflex the metronome is sounded for 30 seconds during which the secretion of saliva is measured in drops, and at the same time the interval between the beginning of the stimulus and the beginning of the salivary secretion is recorded. This interval is customarily called the latent period, although as [p. 49] will be seen later some other term might more usefully have been employed. Stimulation by the metronome is not followed in this particular experiment by feeding, i.e. contrary to our usual routine the conditioned reflex is not reinforced. The stimulus of the metronome is repeated during periods of 30 seconds at intervals of two minutes. The following results are obtained:
The continuation of this experiment must be left over until later on in the lecture when it will be possible to add a further important detail. One detail, however, already stands out quite clearly, namely that repeated application of a conditioned stimulus which is not followed up by reinforcement leads to a weakening of the conditioned reflex. If the experiment had been pushed further there would have come a stage when the reflex would entirely disappear. This phenomenon of a rapid and more or less smoothly progressive weakening of the reflex to a conditioned stimulus which is repeated a number of times without reinforcement may appropriately be termed extinction of conditioned reflexes. Such a term has the advantage that it does not imply any hypothesis as to the exact mechanism by which the phenomenon is brought about.
Abundant evidence has been collected in our laboratories relating to experimental extinction of conditioned reflexes, but before I can discuss this it is necessary to make a few remarks about the terminology which will be employed. Formerly we made a distinction between "natural" and "artificial" conditioned reflexes, "natural" reflexes being those which appeared to be formed spontaneously as a result of the natural association of, for example, the sight and smell of food with the eating of food itself, or of the procedure of introducing acid or some rejectable substance with the acid or the rejectable substance itself, while "artificial" reflexes were those which could be formed as a result of artificially associating [p. 50] with the food or rejectable substance stimuli which in the ordinary with food or the rejectable course of events have nothing in common substance. At the present time, however, we know that there is not the slightest difference in properties between all these reflexes. I mention this fact here because the numerous experiments of the earlier period of our work were carried out with the "natural" conditioned reflexes, and it is from these that I shall draw many examples in the present lecture. All the numerous artificial stimuli which we now use every day in our experiments were important to us at the time of those experiments because they provided easily controlled, exact, and regularly reproducible stimuli, and because they could be applied to check the correctness of our conception of the mechanism by which natural conditioned reflexes are formed. At present the artificial stimuli predominate in importance because of the vast field of research they have unfolded to us and because they came ultimately to provide the most important material for our investigation.
The progress of experimental extinction is often subject to fluctuation. The fluctuations of an otherwise smooth curve may be brought about both by external and internal factors. To obtain a smooth curve of extinction of a conditioned reflex it is necessary to maintain the unreinforced conditioned stimulus rightly constant in character and strength; the environing experimental conditions also must remain absolutely constant. Very wide fluctuations in the reflex undergoing experimental extinction are apt to occur in the case of 8 natural conditioned stimulus, for example the presentation of food, which may be held at one time further away from, the animal than at another, or which may be held stationary or slightly moving. With an artificial conditioned stimulus, on the other hand, it is quite easy to obtain an exact repetition of the stimulus and so to avoid this cause of disturbance in the curve of extinction. With regard to variations in the experimental conditions it is only natural that any marked changes in the environment, such, for example, as any introduction of strong extra stimuli which would produce external inhibition, should also affect the smoothness of the curve of experimental extinction. Such strong stimuli abruptly diminish all conditioned reflexes, including of course reflexes undergoing extinction, but the reflexes reappear when the disturbing stimuli are removed. Even greater interest attaches to the effect of extra stimuli of small intensity: Such stimuli produce a temporary [p. 51] weakening, not of the reflex, but of the progress of the experimental extinction. An example can be seen in the fifth repetition of the conditioned stimulus in our lecture demonstration (p. 49). The rise in the reaction from 5 drops to 7 drops definitely coincided with some small disturbance produced by the audience. This effect of extra stimuli of small intensity is of great importance for the physiology of the hemispheres, and we shall return to it later in this lecture.
Even with stimuli of constant strength, and with constant environing conditions, fluctuations in the curve of experimental extinction are sometimes observed. These fluctuations are of rhythmic character and are evidently due to some internal factors. These factors affect directly the nervous processes involved in experimental extinction, and we shall come across examples on frequent occasions in the further course of our discussion.
The rate of experimental extinction, measured by the period of time during which a given stimulus must be applied at definite regular intervals without reinforcement before the reflex response becomes zero, depends on numerous conditions. First among these come any individual peculiarities of the nervous organization of the animal. Under the same set of external conditions some animals will have the conditioned reflexes rapidly extinguished, while in others the whole process will be much delayed. In excitable dogs the reflexes are mostly slow of extinction, but in quiet animals extinction is rapid. Clearly also the extent to which a reflex has gained a firm footing is an important point: a reflex which has only recently been established-is likely to be less firmly grounded than an older one and is likely to suffer extinction the more quickly. The rapidity of extinction depends also in a great measure upon the intensity of the unconditioned reflex underlying the conditioned one which is undergoing experimental extinction.
In this connection the following experiments by Dr. Babkin are of interest:
An unconditioned reflex to a given quantity of a l% extract of quassia introduced into the dog's mouth produces on an average of ten experiments 1.71 c.c. of salivary secretion. A conditioned reflex established on the basis of this unconditioned one produces 0.3 c.c. during one minute of stimulation. A definite quantity of 0.1% aqueous solution of hydrochloric acid evokes in the same dog an unconditioned reflex measuring on an average of five experiments [p. 52] 52 c.c. The corresponding conditioned reflex gives 0.9 c.c. during one minute of stimulation. The conditioned stimuli are "natural" ones, namely, the presentation of quassia or of acid, as the case may be, at some distance from the animal. Every other condition of the experiment is maintained rigidly constant. The following table illustrates the experimental extinction of each of the two conditioned reflexes in this animal.

Yet another important factor in determining the rate of experimental extinction is the length of pause between successive repetitions of the stimulus without reinforcement. The shorter the pause the more quickly will extinction of the reflex be obtained, and in most cases a smaller number of repetitions will be required. The conditions may be illustrated from an experiment by Dr. Babkin:
The conditioned stimulus was provided by meat powder presented to the dog at a distance for one minute; the stimulus was repeated several times in succession, and was of course not reinforced. The five series of extinctions given below were carried out on the same animal in a single day. Between separate extinctions the dog was given a rest, and the reflex was reinforced by feeding with meat powder.

[p. 53]

The final condition which influences the rate of experimental extinction is the number of times the given reflex has been subjected to extinction in the same animal. After each fresh extinction of a conditioned reflex the number of unreinforced [p. 54] conditioned stimuli required to produce the next experimental extinction is less, until in the end a zero reaction results in some dogs from only a single application of the unreinforced conditioned stimulus.
A circumstance of especial interest is that experimental extinction of any single conditioned reflex results, not only in a weakening of that particular conditioned reflex which is directly subjected to the extinction (primary extinction), but also in a weakening of other conditioned reflexes which were not directly subjected to extinction (secondary extinction). This latter phenomenon involves not only those conditioned reflexes which were based upon a common unconditioned reflex with the primarily extinguished one (homogeneous conditioned reflexes), but also those which were based upon a different unconditioned reflex (heterogeneous conditioned reflexes). Sometimes secondary extinction reaches a profound degree, involving even the unconditioned reflexes. The latter case is illustrated by the following experiment of Dr. Perelzweig:
A given quantity of hydrochloric acid produced on an average in a dog a salivatry secretion of 6 c.c. After several extinctions of the corresponding conditioned defence reflex, in which the conditioned stimulus was tactile, application of the acid itself produced only 3.8 c.c. Even more striking results were obtained in connection with the secondatry extinction of homogeneous conditioned reflexes, and our experiments were mainly concerned with these.
In every case of secondary extinction the degree to which the primary experimental extinction is carried is of first importance. A primary experimental extinction of a conditioned reflex which is carried to its final stages smooths out the many small points of difference between secondary extinctions of different conditioned reflexes, but a primary extinction which is carried to only a moderate degree leaves these small differences well pronounced. It may be stated, other things being equal, that the extent to which homogeneous conditioned reflexes undergo secondary extinction is determined by their relative physiological strengths. The strength of a conditioned reflex, in its turn, depends on whether it has long been established, on the number of times it has been refreshed by reinforcement, and on whether or not the reinforcement has been discontinued, and for how long. The extent of secondary extinction depends also on whether, and on how often, the reflex has previously [p. 55] been subjected to experimental extinction, and on whether or not it is reinforced immediately before the primary extinction is begun.
The greater the strength of the conditioned reflex as compared with the reflex which is subjected to primary extinction, the less does it undergo secondary extinction; on the other hand, if the stronger reflex is subjected to primary experimental extinction the weaker conditioned reflex undergoes complete secondary extinction. The following experiments bearing on this subject were performed by Dr. Babkin :
A dog has three conditioned reflexes to acid, one depending on the sound of a buzzer, a second on the sound of a metronome and a third on a tactile stimulation of the skin. Every conditioned stimulus was continued always for 30 seconds. The first experiment shows the relative strengths of the reflexes.

In the following experiment a primary extinction of the reflex to the metronome was produced by repeating the stimulus at intervals of three minutes.

[p. 56]
On primary extinction of the conditioned reflex of medium strength the weaker tactile reflex also was completely extinguished, while the stronger reflex (to buzzer) was still partly active. These results were corroborated by further experiments in which the order of testing the secondarily extinguished reflexes was reversed.
The same dependence of the degree of extinction upon the strength of the conditioned reflexes is seen when the conditioned reflex undergoing extinction is one to a compound stimulus composed of several distinct elements which can be applied either simultaneously or else independently. Primary extinction of the reflex to the compound stimulus is always accompanied by secondary extinction of the reflexes to its individual components. Supposing there are two components of equal physiological Strength, then the primary extinction of the one leads to simultaneous secondary extinction of the other, while the reflex to the compound stimulus is usually considerably diminished. Where, however, the two components of the compound stimulus are unequal in strength, primary extinction of the stronger reflex leads to a complete extinction of the reflex to the weaker component, while extinction of the weaker (unless carried beyond zero) leads only to a partial weakening of the reflex to the stronger component. The primary extinction of the stronger reflex leads also to a complete extinction of the reflex to the compound stimulus. The question of interrelation between the different individual components in a compound stimulus will be discussed further in a future lecture.
In the meantime we must endeavour to find a correct interpretation of the phenomenon of experimental extinction, and we shall find it important in this connection to give attention to a case in which the weaker of the two components in a conditioned reflex is completely overshadowed by the stronger, the weaker when tested separately producing no positive reflex effect. When in such a case the weaker stimulus is applied singly several times in succession, without reinforcement, there results, nevertheless, an extinction not only of the reflex to the stronger stimulus, but also of the reflex to the compound stimulus. An experiment by Dr. Perelzweig can be taken to illustrate this point:
A compound conditioned reflex has been established on a basis of the defence reflex to acid. The individual components of the compound stimulus are a tactile stimulus and a thermal stimulus of 0° C. In the experiment given below the animal is stimulated by [p. 57] the compound stimulus and by its components separately. In every case the stimulus is applied during one minute.
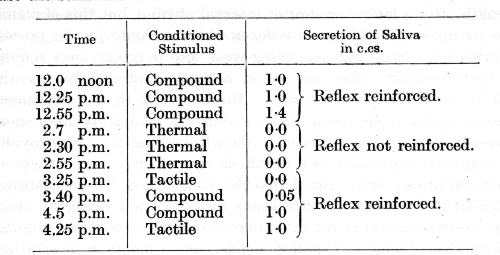
This experiment shows that the application of the thermal component, which by itself was ineffective, led when repeated three times without reinforcement to a complete secondary extinction of the stronger tactile component and to a practically complete extinction of the reflex to the compound stimulus.
Hitherto, when referring to the degree of extinction, we have only spoken of the extinction as being partial or as being complete, but we shall now have to extend our conception. Not only must we speak of partial or of complete extinction of a conditioned reflex, but we must also realize that extinction can proceed beyond the point of reducing a reflex to zero. We cannot therefore judge the degree of extinction only by the magnitude of the reflex or its absence, since there can still be a silent extinction beyond the zero. This statement rests upon the fact that a continued repetition of an extinguished stimulus' beyond the zero of the positive reflex deepens the extinction still further. Such an extension of our conception serves fully to elucidate the experiment just described, and it explains why the seemingly inactive thermal component when subjected to experimental extinction led to such a profound secondary extinction of the stronger tactile component. The importance of considering the degree of extinction in all experiments thus becomes evident. The methods of determining the degree of extinction when it goes beyond zero will be explained in connection with the question which will next be discussed.
We shall consider what happens to the conditioned reflexes after they have been subjected to experimental extinction and inquire [p. 58] whether they ever regain their original strength. Left to themselves extinguished conditioned reflexes spontaneously recover their full strength after a longer or shorter interval of time, but this of course does not apply to conditioned reflexes which are only just in process of formation. Such reflexes, being weak and irregular, may require for their recovery after extinction a fresh reinforcement by the underlying unconditioned reflex. However, all those conditioned reflexes which have been fully established invariably and spontaneously return sooner or later to their full strength. This provides one way of determining the depth of extinction; it is measured, other conditions being equal, by the time taken for spontaneous restoration of the extinguished reflex to its original strength. Such time interval may vary for the different reflexes from a few minutes to a number of hours. I shall give a few experiments in illustration. The first is an experiment by Dr. Babkin:
Presentation of meat powder a short distance away at intervals of three minutes ; the reflex is not reinforced.

The second experiment is by Dr. Eliason :
Meat powder is presented a short distance away at intervals of ten minutes; the reflex is not reinforced.

It will be interesting now to return to the demonstration given in the earlier part of this lecture and to examine whether there is [p. 59] any spontaneous recovery of the reflex to the metronome which was partially extinguished just twenty-three minutes ago.
Continuation of Demonstration.-- The dog is again subjected to the stimulus of the metronome for 30 seconds: the latent period of the conditioned reflex now comes out at 5 seconds and the salivary secretion is 6 drops. At the last reading the latent period was 13 seconds and the salivary secretion 3 drops. Considerable recovery has therefore taken place spontaneously during the lecture.
The great differences in rapidity of restoration of extinguished reflexes depend on a number of factors. The most important factor is the depth of the preceding extinction. The individual character of an animal and its type of general nervous organization also play an important part. Much depends also on the intensity of. The conditioned reflex which was subjected to experimental extinction; and finally, upon how often the experimental extinction has been repeated. In every case, however, it is possible to accelerate the restoration of an extinguished conditioned reflex. For this purpose it is only necessary to apply the unconditioned stimulus on which the conditioned reflex was built up, either singly, or together with the extinguished conditioned stimulus. This method produces a more or less rapid restoration, according as the conditioned reflex has been extinguished to a greater or less degree. If the extinction has not been carried very far, a single application of the unconditioned stimulus is often sufficient to restore the reflex to full strength; but if the extinction has been made profound, repeated reinforcements are necessary. This means of accelerating the recovery of the extinguished reflex affords another method of measuring the depth of extinction. The further question whether the acceleration in the restoration of an extinguished conditioned reflex is greater when the unconditioned reflex is applied singly, or when it is applied in the form of reinforcement cannot be discussed at present, as it is still under investigation (see Lecture XXII).
All this description of facts about extinction may have proved rather wearisome to the reader, owing to the absence of any underlying uniformity. Nevertheless they served an important purpose in that through a careful consideration of them we were enabled gradually to come upon a solution of the fundamental question as to the intimate nature of experimental extinction. By ruling out one interpretation after another we arrived at the conclusion that [p. 60] extinction must be regarded as a special form of inhibition. That it cannot be regarded as an irreparable destruction of the conditioned reflex, due to disruption of the respective nervous connections, is evidenced by the fact that the extinguished reflexes spontaneously regenerate in course of time. Another possible explanation also suggests itself: may it not be that the experimental extinction is brought about simply by fatigue in some part of the neuro-secretory apparatus involved in the reflex? This is ruled out by the following evidence. The secretory elements in the gland do not become fatigued when the conditioned reflexes are being reinforced, although they continue indefinitely during an experiment to produce a full salivary secretion. Moreover, the restoration of an extinguished reflex is greatly accelerated by a fresh application of the unconditioned stimulus, a still further secretory activity of the gland being readily obtained although the reflex has been deeply extinguished. Neither can there be fatigue in the nervous centres of the secretory reflex. It is sufficient to recall the experiment which the conditioned reflex to the compound stimulus which had two cutaneous components -- tactile and thermal. The thermal stimulus, which was the weaker, could not by itself produce even the slightest positive effect: yet none the less its repeated application brought about a secondary extinction of the stronger tactile stimulus and even of the compound stimulus itself. The extinction of the ineffective thermal stimulus was at no time accompanied by any kind of positive activity of the nervous elements, and it is difficult to conceive that a part of the central nervous system underwent fatigue without previous activity. Again, it would seem that if we were to admit the possibility of any fatigue in the nervous centres, we could expect only the reflex to the thermal stimulus to become fatigued, but we find also an extinction of the reflex to the tactile stimulus which was not brought into activity at any time during the repeated stimulation of the thermal receptors. Thus by a process of elimination we are forced to the conclusion that experimental extinction is based on inhibition, and if we look at the facts which have been described, in the light of this conclusion nearly all of them become perfectly intelligible.
The spontaneous rhythmic fluctuations in the reflexes sometimes observed during the process of experimental extinction can now easily be explained as a manifestation of the struggle which is taking place between the nervous processes of excitation and inhibition [p. 61] before one or other of them gains the mastery. Similarly it becomes quite easy to understand the part played by the individuality of the animal. We have all observed for ourselves how the inhibitory processes in the nervous system of human beings are seldom of the same intensity in any two people, and numerous examples in the further course of these lectures will make it clear that a precisely similar variation obtains in the nervous system of animals.
It is clear that the more vigorous a conditioned reflex, or in other words the greater the intensity of the excitatory process, the more intense must be the inhibitory process in order to overcome it, and therefore the greater the number of unreinforced repetitions necessary to bring about complete extinction. Again, it was seen that a repetition of the non-reinforced conditioned stimulus was necessary to produce a sufficient summation of the inhibitory after-effect for complete experimental extinction, and it is reasonable to suppose that the shorter the intervals between successive repetitions of the stimulus the more quickly will the required intensity of the inhibitory process be obtained. This also was found to be the case. As a result of repetitions of experimental extinction on the same animal the zero level of a fresh extinction of the reflex is reached more rapidly. This shows that inhibition like excitation is facilitated by repetition. The fact itself is well known from observation of ourselves and others, but abundant experimental evidence for animals will be afforded during the further course of our study of conditioned reflexes.
The influence exerted by experimental extinction on reflexes other than the one undergoing extinction, including unconditioned reflexes as well as homogeneous and heterogeneous conditioned reflexes, must be regarded as the result of a spreading of the inhibitory process from its point of initiation through the entire nervous structure of the hemispheres. This process will be fully discussed in one of the later lectures.
We have now to consider in detail still another important feature which has already been noted in passing, but was left unexplained, namely the frequent deviations observed in the curve of experimental extinction. These deviations represent sudden rapid strengthenings in the intensity of the reflex which is undergoing extinction, and they depend on the introduction of any accidental stimuli into the experimental environment. Some extraneous sound or shadow [p. 62] finding its way into the room produces at once a rapid strengthening of the reflex, and of course a similar effect is produced by different extra stimuli which we ourselves apply on purpose in order to study this phenomenon experimentally.
I shall describe first of all an observation which for a long time we were at a, loss to interpret. A natural conditioned reflex to meat powder, which, as we know from control experiments, after extinction recovers its initial value spontaneously in something between a, half and one hour, is again extinguished to zero. This time, however, instead of waiting for the spontaneous recovery of the reflex a weak solution of acid is immediately introduced into the dog's mouth, and after the termination of the secretion produced by the acid (about five minutes) meat powder is again presented at a' short distance. This time although nothing like half an hour has elapsed the conditioned alimentary reflex is found to be almost completely restored. At first sight the accelerated recovery of the extinguished reflex seems paradoxical, since we know already that positive conditioned reflexes are always quite definitely specific -- a definite stimulus rigidly evoking a definite reaction -- but in this case a stimulus to an extinguished conditioned alimentary reflex has had its full strength restored through the single application of a stimulus to a heterogeneous unconditioned reflex, namely the defence reflex to acid. And there can be no doubt that although the secretory component of the two reflexes is effected through the same glands yet they are distinctly heterogeneous in nature, since the defence reflex to acid differs sharply from the alimentary reflex to food both as regards the composition of the saliva secreted and as regards the character of the motor response. Without attempting for the present to give any explanation we can designate this observation from a purely matter of fact point of view as consisting of a sudden removal by an extraneous reflex of the inhibitory process set up by experimental extinction.
The whole group of cases of which the above is an illustration have one common feature. In all of them the removal of the inhibition is only temporary, persisting no longer than the extra stimulus responsible for the removal of inhibition and its after-effect.
It is interesting to mention in this connection a disagreement which arose among the members of the staff in our laboratory before the fact of the restorative effect of acid upon an extinguished [p. 63] alimentary conditioned reflex had been indubitably established. Some of the workers admitted this restorative effect without question, while others disputed it. However, the experimental side of the question turned out to be right in both cases. The cause of the discrepancy was clearly brought out in Dr, Zavadsky's researches. It appeared that previous observers had overlooked the fact that their conditions of experimentation were not fully identical. Those workers who accepted the restorative effect had tested the extinguished conditioned alimentary reflex immediately or only a few moments after the salivary secretion in response to the acid had ceased, while the others had tested the extinguished reflex after allowing a considerable interval of time to elapse. Realizing the difference in the experimental procedure of the two sets of workers Dr. Zavadsky was able in his experiments to obtain all the different stages that had been reported by other workers. Two of his experiments, performed on the same day, are given below.
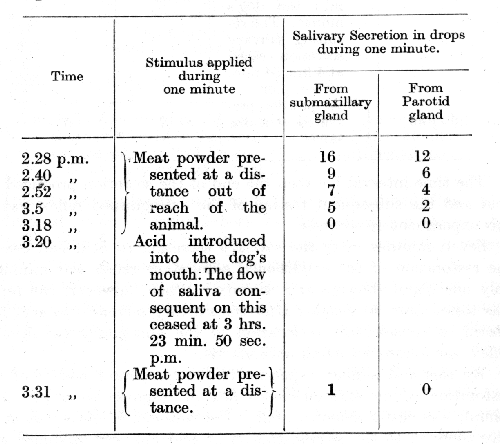
The time interval between the end of the secretion produced by acid and the subsequent testing of the extinguished reflex was in the above experiment 7 minutes 10 seconds.
[p. 64]
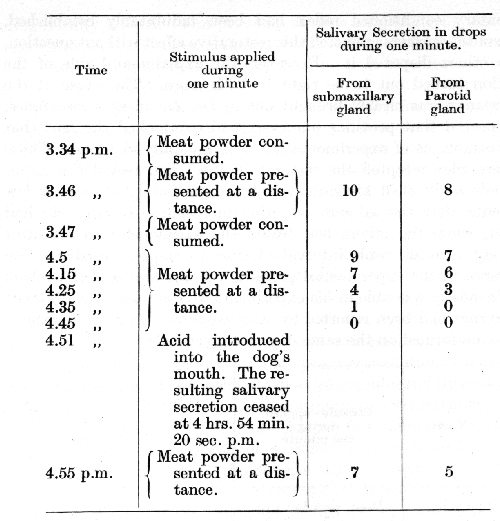
The time interval between the end of the secretion produced by acid and the subsequent testing of the extinguished reflex was in this experiment 40 seconds.
Seven minutes after the salivary secretion to acid had ceased the restoration of the conditioned alimentary reflex was minimal, only one gland showing any activity. When, however, the reflex was tested only 40 seconds after the salivary secretion to acid had ceased, a considerable restoration of the alimentary conditioned reflex was found, involving both glands.
By this and similar experiments the temporary nature of the restoration of an extinguished reflex in response to other extra stimuli was easily demonstrated. The restorative effect was in no way confined to the administration of acid but was produced also by any other extra stimulus. A further example from Dr. Zavadsky's experiments illustrates this general case. The experiment was conducted on another dog.
[p. 65]
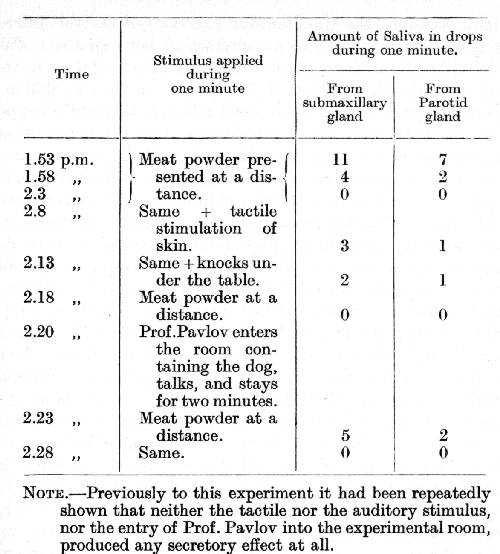
This experiment leaves no doubt that the extinguished alimentary conditioned reflex is restored both by the actual presence of the extra stimulus (tactile and auditory), and by its after-effect (after-effect of stimulus of my entering the room).
In all the experiments which have just been described the restoration of the extinguished reflexes lasts only for a few minutes, depending on the duration of the extra stimulus and its after-effect. In the case, however, of certain special extra stimuli already mentioned in connection with external inhibition, stimuli which are of a protracted nature, the restorative effect is felt throughout the whole course of experimental extinction, which is therefore never smoothly progressive and can never be brought down to and kept at the zero level of the reflex.
We have now to discuss another important observation bearing on the same point. During the whole period of our work we observed [p. 66] on many occasions the simultaneous existence of several different reflexes, leading of course to an interaction between them which resulted either in predominance of one or another reflex or in their mutual neutralization. Thus, if we make a tactile stimulation of the skin the stimulus to a conditioned reflex, it frequently happens that we are bothered with an interference from the unconditioned reflex response to the cutaneous stimulus itself, in the form of the scratch reflex or some sort of quivering reflex. This may, in rare cases, be so troublesome that the conditioned reflex never reaches a stable value. Exactly the same thing happens sometimes with musical tones of exceedingly high pitch, it being in some dogs impossible to overcome the difficulty of the resulting sharp motor response. All such powerful unconditioned stimuli exercise an external inhibitory influence which perpetually interferes with all positive conditioned reflexes. But it is obvious that these persistent extra reflexes should exert a still more powerful disturbing influence upon the normal course of the inhibitory processes underlying extinction, since inhibition is in every respect more labile than excitation. I shall be giving a number of examples substantiating this statement in a further lecture, when the whole matter will be subjected to a rigorous experimental analysis. All the considerations put forward in this lecture permit us to regard the temporary restoration of the reflex which is in process of extinction, or which is already extinguished, as based upon the removal of an inhibitory process. We therefore describe this phenomenon as a dis-inhibition, a term we shall always use in the future when we wish to denote a temporary removal of inhibition.
The next question is, whether any distinction can be drawn between the ease of the restoration of an extinguished conditioned reflex resulting from fresh applications of the appropriate unconditioned stimulus, and the case which has just been termed dis-inhibition. Our experiments show that undoubtedly such a distinction does exist. In the first case, when restoration is effected by the special unconditioned stimulus underlying the reflex which has undergone extinction, such restoration is permanent. In the second case, however, when the restoration is effected under the influence of any alien stimulus, such restoration is only temporary. As to the actual reason for this difference it is not possible to say very much upon the experimental evidence available up to the present. There is, however, no doubt that in the first case, just as has already [p. 67] been shown in the second, we are dealing with a removal of inhibition. Any hypothesis of an irreparable destruction of the conditioned reflex in the process of experimental extinction cannot possibly stand for a moment, since in every case of extinction the reflex invariably becomes spontaneously restored in a longer or shorter time.
The question of the difference in the mode of restoration in these two cases probably goes much deeper, involving the intimate nature of the nervous process underlying dis-inhibition. Regarding the nervous mechanism of dis-inhibition we cannot hope at present to approach anything like a fundamental conception, since as yet we know little about the real nature either of the inhibitory process, or of the excitatory process, or of their mutual relations.
I should like, however, specially to direct your attention to one very important feature which repeatedly enforces notice. We have seen that the very same extra stimuli, which, when they evoke strong extraneous reflexes, produce external inhibition of the positive conditioned reflexes, produce, when their effect is weak from the start or weakened by repetition, dis-inhibition of the conditioned reflexes which were made to undergo extinction. Many examples of this will appear in the next lecture. We are now afforded some justification for regarding dis-inhibition, as we did a short while ago, as being the " inhibition of an inhibition." By this we do not pretend, however, to explain the underlying mechanism of dis-inhibition.
The main conclusion of our discussion of the experimental evidence described in this lecture can be summed up briefly as follows. A stimulus to a positive conditioned reflex can under certain definite conditions readily be transformed into a stimulus for a negative or inhibitory conditioned reflex; this transformation is fairly rapid, smooth and progressive. It becomes obvious therefore that in our further study of the function of the cerebral hemispheres we shall necessarily be dealing not only with positive but also with negative or inhibitory conditioned reflexes.