
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE V
Internal inhibition (continued): (b) Conditioned Inhibition.
The fourth lecture was devoted entirely to the study of the first type of internal inhibition, which was termed experimental extinction. In extinction the positive conditioned stimulus is temporarily transformed into a negative or inhibitory one by the simple method of repeating it several times in succession without reinforcement. In the present lecture we shall consider the second type of internal inhibition, which has also been investigated in some detail.
The method of experimentation is as follows. A positive conditioned stimulus is firmly established in a dog by means of the usual repetitions with reinforcement. A new stimulus is now occasionally added, and whenever the combination is applied, which may be at intervals sometimes extending to hours or days, it is never accompanied by the unconditioned stimulus. In this way the combination is gradually rendered ineffective, so that the conditioned stimulus when applied in combination with the additional stimulus loses its positive effect, although when applied singly and with constant reinforcement it retains its full powers.
We have been accustomed in our investigations to designate this phenomenon by the name of conditioned inhibition, although this cannot be regarded as especially appropriate since the development of experimental extinction also is subject to equally rigid conditions. The use of this term, in fact, can only be justified by historical consideration. Since we were concerned in this case with the participation of an additional stimulus, the whole phenomenon was confused at first with external inhibition. It was only later when its character of internal, as distinct from external, inhibition became firmly established that the prefix "conditioned" was added. As will be shown further on, this form of inhibition might more appropriately have been termed "differential inhibition."
The process of development of conditioned inhibition presents [p. 69] especial interest. While illustrating the varied complexity of the phenomena involved, it demonstrates at the same time the value of the experimental method as providing a satisfactory means for the analysis of this very complexity into simple general principles. For this reason we shall discuss conditioned inhibition in considerable detail.
The first point of importance in the establishment of a conditioned inhibition is its dependence on time relations between the applications of the two stimuli in the inhibitory combination. Conditioned inhibition is developed with comparative ease in all those cases where the duration of the positive stimulus overlaps that of the additional stimulus. In our experiments it is usual to start the additional stimulus a few seconds (generally from 3 to 5) before the positive stimulus, but provided there is an overlap it is immaterial whether the commencement of the additional stimulus precedes, coincides with, or even follows by a few seconds, that of the positive stimulus. If, on the other hand, the additional stimulus is removed as soon as the positive stimulus is applied, so that the two stimuli never coincide, the development of the conditioned inhibition may be a, matter of considerable difficulty, and accompanied by restlessness and various defence reactions of the animal. If, finally, a pause of several seconds is introduced between the termination of the additional stimulus and the beginning of the positive stimulus no inhibition develops at all. On the contrary, in the majority of cases, when this pause reaches a duration of about ten seconds, the additional stimulus itself acquires the properties of a positive stimulus. This has been discussed already as the general method of formation of positive conditioned reflexes of the second order. It is only with exceptionally powerful additional stimuli, such for example as a powerful motor-car hooter, that the pause can be increased to so much as twenty seconds and a conditioned inhibition still be developed. An example from the work of Dr. Frolov will serve to illustrate the latter case :
A motor-car hooter was allowed to act for 10 seconds, when, after a pause of a further 10 seconds, the alimentary conditioned stimulus of the metronome was applied. The first application of the hooter did not in the least diminish the magnitude of the succeeding reflex reaction. When, however, the same combination had been repeated several times in succession, and always without reinforcement, the reflex began gradually to diminish. [p. 70] This diminution persisted even when the pause was lengthened to 20 seconds.
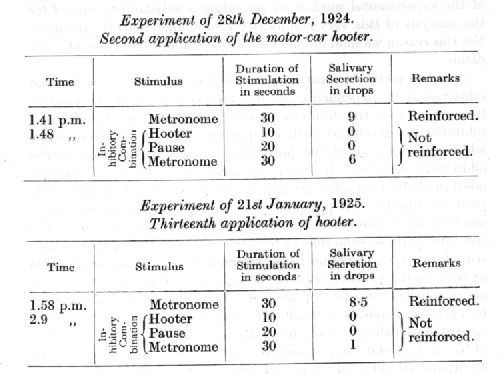
We thus find that the time interval between the two stimuli required to produce either conditioned inhibition, or alternatively a positive conditioned reflex of the second order, varies according to the intensity of the additional stimulus. This contingency of the ultimate significance of the additional stimulus upon the time relations is interesting as evidence of an encounter between the antagonistic processes of excitation and inhibition.
The following interpretation of the complete difference in the final character assumed by the additional stimulus under so slight a difference in the experimental conditions seems best to agree with the experimental results : When the additional stimulus or its fresh trace loft in the hemispheres coincides with the action of the positive stimulus, there must result some sort of special physiological fusion of the effect of the stimuli into one compound excitation partly differing from and partly resembling the positive one. It will be shown in the seventh lecture, that in response to closely related stimuli, such as neighbouring tones or tactile stimulation of adjacent [p. 71] places of the skin, the same events take place as in the development of conditioned inhibition. If one stimulus with the help of the usual procedure is given positive conditioned properties, the neighbouring stimuli belonging to the group also give at first a positive conditioned reaction, whereas later, on their systematic repetition without reinforcement, they lose their excitatory properties and acquire inhibitory properties instead. This result corresponds exactly with the successful development of conditioned inhibition in the experiment mentioned at the beginning of this lecture. When, however, the additional stimulus is separated from the application of the positive conditioned stimulus by a longer interval, the union in the hemispheres into a single compound excitation does not materialize; instead, the stimuli act upon the hemispheres as two distinctly separate events, and the additional stimulus acquires new properties as a positive conditioned stimulus of the second order. The older conditioned stimulus acts in this case in exactly the same capacity as an unconditioned stimulus in the usual method of establishing conditioned reflexes of the first order. A strong stimulus has a prolonged after-effect; it is, therefore, still capable of being fused by the hemispheres with the conditioned stimulus into a special new compound even after a long interval of time. It is easy to understand from this point of view that in the establishment of conditioned inhibition the interval which can be made between the end of the additional stimulus and the beginning of the positive conditioned stimulus must depend directly upon the strength of the additional stimulus. Whether our interpretation be the correct one or not, the phenomena are encouraging to the experimenter in that in every case he can discern a remarkable regularity in these very complex activities of the cortex.
In addition to the usual results obtained as described above, mention must be made of certain very rare phenomena obtained with some apparently normal animals and with animals in which surgical interference with the hemispheres had produced a condition of obviously increased excitability of the nervous system. In these cases even perfectly synchronous applications of the additional and of the positive conditioned stimuli led not to conditioned inhibition but to the development of a definite positive conditioned reflex of the second order. This generally persisted for a very long time, but later ft often happened that the secondary conditioned reflex and also conditioned inhibition were both present simultaneously. [p. 72]
Such a case is represented in the following experiment of Dr. Kasherininova :
Tactile stimulation of the skin was used in this dog as the conditioned stimulus for the defence reflex to acid, and a metronome served for the additional stimulus in an inhibitory combination which, of course, was never reinforced. The twenty-fifth application of the inhibitory combination evoked a secretion of only 3 drops of saliva during one minute, whereas the conditioned stimulus applied singly evoked a secretion of 29 drops during one minute. After thirty-four repetitions of the inhibitory combination the stimulus of the metronome applied singly elicited a salivary secretion of 8 drops, although prior to its participation in the inhibitory combination it had no excitatory effect whatsoever. It is clear, therefore, that the metronome coupled with the tactile stimulus exerted a strong inhibitory influence, but it acquired at the same time some of the excitatory properties of the tactile stimulus, so that when used alone it behaved as a conditioned stimulus of the second order. Though cases such as that just described have been observed but rarely, it is quite possible that a transitory acquisition of weak excitatory properties of the second order is of no infrequent occurrence in the development of conditioned inhibition.
The course of development of conditioned inhibition is not always the same. In some cases the first addition of a new stimulus to the positive conditioned stimulus immediately results in a diminution, or even in a complete disappearance, of the conditioned reflex. With successive repetitions of the combination the reflex comes back nearly to its original level and then again falls slowly to zero. In other cases the first few combinations result in an augmentation of the reflex as compared with the normal isolated action of the conditioned stimulus, and only subsequently does the reflex gradually diminish to zero. In yet a third group it is found that an initial diminution in the strength of the reflex is followed by a phase of augmentation above normal and then again the reflex slowly falls to a permanent zero. All these different phases which are observed during the establishment of the conditioned inhibition depend entirely upon the intensity of the extraneous reflex which is evoked in the animal by the additional stimulus. The initial diminution of the reflex which occurs in one group during the fist few applications of the combination is due undoubtedly to external inhibition. When the additional stimulus applied singly evokes in the dog a [p.73] strong investigatory reflex it is found that its addition to the positive conditioned stimulus exerts an inhibitory influence from the very start. When on the other hand the stimulus evokes only a mild investigatory reflex, the preliminary phase of diminution of the reflex is absent and the first applications of the combination produce an increase in the reflex. This increase is undoubtedly due to dis-inhibition, since -- as we may state now in anticipation of a future discussion -- the positive effect of the majority of conditioned stimuli in our experimental reflexes is almost invariably preceded by a phase of internal inhibition. The inhibitory process is removed by the investigatory reflex, provided that the latter is not sufficiently strong also to influence the excitatory component of the conditioned reflex. The third group of cases, in which the initial diminution in strength of the reflex is followed by an increase above normal, depends on the gradual weakening through repetition of the extraneous reflex produced by the additional stimulus. The external inhibition brought about by the investigatory reaction is at first strong enough to inhibit the conditioned reflex, but, on weakening, produces only dis-inhibition and so increases the strength of the reflex response. Several examples of this will be given at the end of the present lecture.
It is obvious that any agent in nature may be used as a stimulus for the development of a conditioned inhibition, supposing of course that the organism is provided with the requisite organs for the perception of such an agent. The records of actual experiments will afford numerous illustrations of the different types of stimuli employed.
As was mentioned before, not only the actual stimulus but also its trace in the hemispheres can be used for the development of conditioned inhibition. The trace must, however, in all cases be as recent as possible. The establishment of conditioned inhibition when the interval between the stimuli is more prolonged can be brought about only by the use of exceptionally strong additional stimuli. Once, however, the inhibitory combination has been firmly established the pause between the end of the additional stimulus and the beginning of the positive conditioned stimulus may be extended even to so much as one minute without the inhibitory effect of the combination being impaired.
Some experiments have been made in which the time interval was itself employed as a stimulus for internal inhibition. [p. 74]
The following experiment by Dr. Krjyshkovski illustrates such cases:
A definite tone served as a positive conditioned stimulus to acid, while a tactile stimulation of the skin served for the additional stimulus in the inhibitory combination. The inhibitory combination in this dog had been habitually applied for some other special purpose at the 19th-20th minute after the last introduction of acid. This led to the result that at the 19th-20th minute only a minimal salivary secretion was produced by the action of the positive conditioned stimulus when it was applied singly.

In these experiments the interval of 19 to 20 minutes had itself acquired sufficient inhibitory properties to abolish or greatly reduce the effect of the conditioned stimulus.
The rate of development of conditioned inhibition as well as its completeness (absolute or relative inhibition) also depends upon a number of conditions. Of first importance in this connection is the individuality of the animal, the excitable or inhabitable character of its nervous organization. In some dogs the establishment of a conditioned inhibition takes a long time and never becomes absolute; in other dogs an inhibition becomes completely and firmly established after very few repetitions of the inhibitory combination.
A further important factor is the intensity of the additional stimulus employed in the inhibitory combination. For example, in an experiment by Dr. Mishtovt, a metronome was used for a positive conditioned stimulus to acid, while a thermal stimulation of the skin was employed as the additional stimulus. It was found that with the thermal stimulus at a temperature of 4 to 5° C. the first indication of a conditioned inhibition could be observed only after the 30th application of the inhibitory combination, and the inhibition was not yet complete even after 145 applications. With the use, however, of a thermal stimulus at a temperature of 1° C., in an ex-[p.75]periment performed on the same animal after an interval of four months, only twelve repetitions of the inhibitory combination were required to establish complete inhibition.
The rate of formation of conditioned inhibition depends, again, on the character and the relative intensity of the additional stimulus in comparison with the conditioned stimulus. In some dogs it was found impossible to establish complete inhibition by the addition of a thermal stimulus at 45° C., when the sound of a metronome served as the positive conditioned stimulus. When, however, a visual stimulus was employed as the positive conditioned stimulus in a similar combination, a conditioned inhibition could be readily produced [experiment by Dr. Poursikov].
Finally, it should be mentioned that, although other factors may remain constant, the first establishment of a conditioned inhibition in a dog takes more time than any succeeding one.
I have not up to the present given any conclusive evidence that the phenomenon of conditioned inhibition is really in the nature of an inhibition at all, and is not merely a passive disappearance of the positive conditioned reflex owing to the compound stimulus remaining habitually unreinforced. The proof that the phenomenon actually does represent a real inhibition will be gradually brought out as we proceed with our study of the experimental evidence.
It will be of interest, first of all, to study the nature and ultimate function of the additional stimulus on which conditioned inhibition depends. This can, of course, be determined only by trying out the action of the additional stimulus in different modifications of the experiment: Tested singly after the conditioned inhibition has been fully established it produces no positive effect at all. The action of the additional stimulus can be tested, however, by applying it in combination with some other positive conditioned stimulus with which it has never previously been associated. In such a case the inhibitory properties of the additional stimulus become clearly revealed, the result being an immediate diminution in the positive reflex response. This is true not only in the case of homogeneous reflexes, but also in the case of heterogeneous reflexes, and the inhibitory effect may extend even to the unconditioned reflexes themselves. These facts are clearly exhibited in the following experiment by Dr. Leporsky:
Three alimentary Conditioned reflexes have been established in the dog used for this experiment, the three stimuli being the flash [p. 76] of a lamp, a rotating object, and the tone C sharp of a pneumatic tuning fork. Two independent conditioned inhibitions of the reflex to rotation have also been firmly established, one by the use of tactile stimulation of the skin and the other by the use of a metronome.
In the first experiment the flash of the lamp is for the first time accompanied by a tactile stimulation of the skin:
In the second experiment the tone C sharp is for the first time accompanied by the metronome.

It will be seen that the additional stimulus when applied for the first time in the new combination produced a diminution almost to zero in the conditioned reflex response. It follows, therefore, that when an additional stimulus is used with an alien homogeneous conditioned reflex its inhibitory property becomes thereby immediately revealed. This same property is seen also when the additional [p. 77] stimulus is combined for the first time with a heterogeneous conditioned reflex, a case which is illustrated in the following experiment:
A dog has a conditioned alimentary reflex which has been established by the use of the metronome, while the addition of a whistle provides a powerful inhibitory combination. Besides this a, conditioned reflex to acid has been established in response to tactile stimulation of the skin. The metronome and the tactile stimuli belong therefore to heterogeneous conditioned reflexes, and the positive effect of one of them (i.e. the metronome) is completely inhibited by the sound of the whistle. The whistle is now for the first time combined with the heterogeneous tactile conditioned stimulus [experiment by Dr. Babkin].
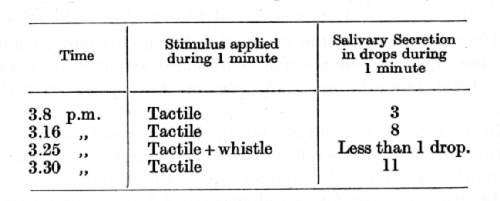
In all the foregoing experiments it had, of course, to be shown before any particular additional stimulus was used in combination with an alien positive conditioned stimulus that it would not exercise any effect of external inhibition. With this precaution it seems to me that the experiments justify our conclusion that where a conditioned inhibition has been firmly established the additional stimulus itself acquires inhibitory properties which can be manifested outside the parent combination. The additional stimulus is therefore termed in our investigations the conditioned inhibitor.
It is made clear by further experiments that a real inhibitory effect is also produced by the inhibitory combination itself, the inhibitory process persisting as an after-effect which may be detected some considerable time after the stimulus of the inhibitory combination itself has been removed. The inhibitory after-effect exerts its influence not only upon the particular reflex to the conditioned stimulus employed in the combination, but also upon all other conditioned reflexes whether homogeneous or heterogeneous. In this connection we may consider the following experiments: [p. 78]
In the first experiment a rotating object serves as an alimentary conditioned stimulus, and a tone of 30,000 vibrations produced by a Galton's whistle as its conditioned inhibitor [experiment by Dr. Nikolaev].

After withdrawal of the stimulation by the inhibitory combination the effect of the positive conditioned stimulus is weakened for several minutes, and only regains its normal strength by degrees.
In the next experiment a rotating object serves as a conditioned stimulus for the defence reflex to acid, and a musical tone serves as a conditioned alimentary stimulus. A tactile stimulus is used as a conditioned inhibitor for the alimentary reflex. [Experiment by Dr. Ponisovsky.]

This experiment demonstrates that the heterogeneous conditioned reflex also becomes diminished as a result of the inhibitory after-effect of the combination.
When the inhibitor is applied by itself alone, and not together with the excitatory stimulus in combination with which it was originally developed, it can also be demonstrated to produce an inhibitory after-effect. [p. 79]
The degree to which positive conditioned reflexes are influenced by the conditioned inhibitor varies inversely as their relative physiological strength. An experiment may be taken from a paper by Dr. Leporsky in illustration of this point :
Three independent conditioned alimentary reflexes have been firmly established to a rotating object, to the flash of several electric lamps, and to a musical tone. A tactile stimulation of the skin has been established as a conditioned inhibitor for each of the three reflexes, so that in combination with any one of them separately it reduces the reflex to zero. All three positive stimuli applied together produce a much greater salivary secretion than any one of them applied singly, showing summation of conditioned reflexes. The experiment shows the effect of the application of the conditioned inhibitor in conjunction with the simultaneous action of all three positive stimuli.

The conditioned inhibitor, therefore, although it reduced to zero every one of the positive conditioned reflexes taken singly, could only partially inhibit the reflex secretion evoked by all three acting simultaneously (2.27 p.m.).
Two further details concerning the inhibitory after-effect remain to be considered. The first is the phenomenon of summation of after-effect. If the inhibitory combination is applied not once only, but several times in succession, so much the more will the strength and duration of the inhibitory after-effect be increased. Two experiments of Dr. Chebotareva, carried out on successive days, serve to illustrate the case in point:
A metronome serves as a conditioned alimentary stimulus, and a rotating object as its conditioned inhibitor. [p. 80]
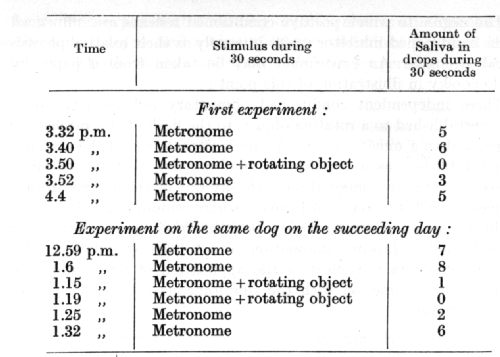
In the first experiment it is seen that the conditioned reflex, which, to start with, was of considerable strength, when tested by a fresh application of the positive conditioned stimulus 1 1/2 minutes after the cessation of a single application of the inhibitory combination (3.52 p.m.), is diminished by half (i.e. from 6 drops to 3). In the second experiment, taking place on the following day, the reflex, when tested 5 1/2 minutes after the second of two successive applications of the inhibitory combination (1.25 p.m.), is diminished by three-quarters (i.e. from 8 drops to 2).
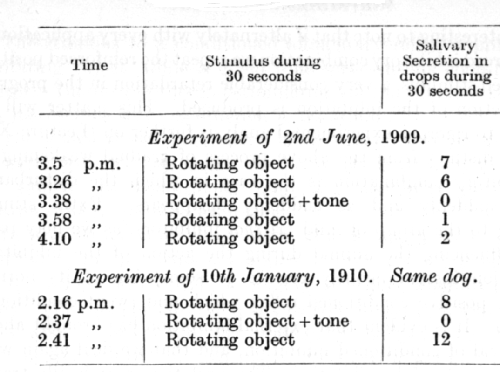
The second important detail concerning the inhibitory after-effect is that its duration becomes shorter as the experiments proceed from day to day. It may extend in the earlier experiments for something over an hour, but in the course of succeeding experiments gradually becomes reduced to only a few minutes or seconds. The following two experiments of Dr. Nikolaev which were performed, with an interval of over 6 months' continuous experimentation, provide an illustration of the case in question:
A rotating object provides the stimulus to a conditioned alimentary reflex, while a given tone serves as its conditioned inhibitor. In the first experiment the inhibitory after-effect was obvious for over 20 minutes; six months later, after continual practice, it was absent so soon as 3 1/2 minutes after the single application of the inhibitory combination.
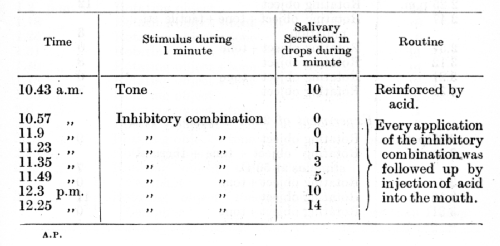
The problem of experimental destruction of the inhibitory properties of the conditioned inhibitor applied singly or in its inhibitory combination is very complex and has not yet been fully worked out. I shall mention, therefore, only some of the better established facts. It is obvious that a complete abolition of the inhibitory properties of the combination should most readily be brought about by reversing the technique employed in its formation -- i.e. by systematically reinforcing the inhibitory combination by the appropriate unconditioned reflex. An experiment by Dr. Krjishkovski will illustrate this process:
A given tone of a pneumatic tuning fork provides a conditioned stimulus to acid, while a tactile stimulation of the skin serves as the conditioned inhibitor.
[p. 82]
It is interesting to note that if alternately with every application of the reinforced inhibitory combinations we repeat the reinforced positive conditioned stimulus, a very considerable retardation in the progress of destruction of the inhibition is produced. This matter will be subjected to rigorous experimental analysis further on (Lecture XI).
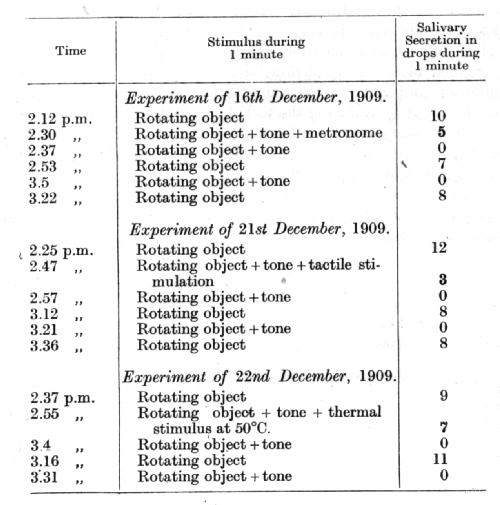
Quite distinct from the above process of gradual weakening of the inhibitory combination is the case in which the disturbance appears suddenly and as suddenly disappears. Extra stimuli belonging to the group of mild external inhibitors, as we may term them, influencing the animal during the action of the inhibitory combination instantaneously restore to something of its normal value the positive conditioned reflex which underwent conditioned inhibition. It is evident that external inhibition has brought about the removal of conditioned inhibition, and that we deal again with the phenomenon of dis-inhibition. These relations may be illustrated by the following experiments of Dr. Nikolaev:
[p. 83]
The stimulus to s conditioned alimentary reflex is provided by a rotating object, while a given tone serves as its conditioned inhibitor. Tactile and thermal stimulations of the skin and the sounds of a metronome serve as different extra stimuli.
The above three experiments show that during the time when the extra stimuli (metronome, tactile, or thermal) were acting upon the animal the conditioned inhibition was partially removed, revealing the underlying excitation.
In connection with these observations considerable interest is attached to the following experiment on the same dog, in which it was the intention to introduce an odour as still another dis-inhibiting agent. For this purpose the dog was transferred into another room fitted with a special box for graduating the intensity of odours. The apparatus itself, in addition to its visual effect and the sound of its electric motor, acted upon the animal by blowing a continuous current of air. All these agencies introduced a whole complex of new extra stimuli into the experiment, even without the addition of the odour. The new complex of extra stimuli dis-inhibited the inhibitory combination, but this effect gradually declined as time went on, disappearing completely in an hour and a half after the experiment was started. The following experiment was performed on the day following the last experiment.
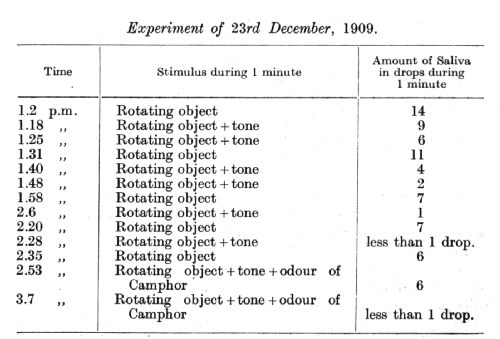
[p. 84]
The first application of the camphor dis-inhibited the combination, while on the second application this extra stimulus had already lost its dis-inhibiting effect. This rapid disappearance of the effect of the extra stimuli introduced by the new apparatus, and again by the use of the camphor, is the most usual case with extra stimuli which bring about dis-inhibition. The dog employed in this particular experiment was an old laboratory animal and had previously been subjected to numerous extraneous agencies, so that different changes produced now only a transient effect, the animal speedily becoming indifferent to them. This is why the introduction of new conditions into this experiment produced a dis-inhibition right from the very start and never any inhibition of tho positive conditioned reflexes.
Results were quite different in the case of another dog used by Dr. Nikolaev. This dog was fresh to the laboratory and may have possessed also a type of nervous organization which was more easily subjected to inhibition. In this dog a rotating object was used for a conditioned alimentary stimulus, and a tone for its conditioned inhibitor. A metronome provided the extra stimulus.

The experiment on the following day gave similar results. On the 18th February, at the beginning of the next experiment, the metronome was applied alone during one minute, the subsequent course of the experiment being shown on the opposite page.
In the experiment of 15th February the sound of the metronome, which served as an extra stimulus, did not produce during its first application the result which would be expected from dis-inhibition (11.41 a.m.). But the very same stimulus of the metronome when [p. 85] applied along with the positive conditioned stimulus used singly produced a complete inhibition (12.14 p.m.). It follows, therefore, that the zero value of the reflex in the first case -- inhibitory combination + metronome -- was not really due to the internal inhibition remaining undisturbed but was due to external inhibition resulting from a very powerful extraneous reflex in response to the first application of the metronome. As a result of several repetitions of the stimulus of the metronome with the inhibitory combination on the 16th February and an application at the beginning of the experiment of 18th February, the stimulus of the metronome was no longer able to produce so powerful an alien reflex and therefore its inhibitory

effect upon the positive conditioned component had practically disappeared (12.27 p.m.). However, its dis-inhibitory effect was still retained and could be well seen in the experiment of 18th February (11.32 a.m. and 12.9 p.m.).
The terminology used in the interpretation of the foregoing phenomena may seem to be artificial and arbitrary, but it is not possible at a time: when the intimate mechanism is still beyond our powers of analysis to avoid such terms in what is necessarily a schematic representation of the complex nervous processes involved. Our terms serve to describe only the actual state and succession of events.
A dis-inhibition of the inhibitory combination can further be obtained by means of those stimuli which determine a permanent external inhibition (p. 46). The following experiment by Dr. Nikolaev bears out this statement: [p. 85]
A rotating object serves for the conditioned alimentary stimulus, while a tone serves for its conditioned inhibitor.

In this case the inhibitory after-effect of sodium carbonate, given 4 minutes before, caused a definite dis-inhibition of the reflex (2.44 p.m.).
If an inhibitory combination is applied early within the time of the after-effect left by a very strong extra stimulus, for example a concentrated solution of quinine, no dis-inhibition is obtained. This corresponds exactly with the case of the metronome in the experiment on page 84· However, the dis-inhibiting effect of quinine can be exhibited as clearly as in the case of the metronome, simply by throwing in the inhibitory combination' at a later stage, when the strength of the after-effect of the action of the; quinine has had time to diminish. On account of the complete accord of the two sets of experiments the description of the experiments with quinine need not be given.
All the experimental evidence which has been dealt with in this lecture establishes conclusively that the nervous processes on which conditioned inhibition depends are identical in character with those of extinctive inhibition. The fundamental condition for their development is the same, namely that there shall be no reinforcement by the unconditioned reflex. In both cases the process develops gradually, being strengthened by repetition. Moreover, the inhibitory after-effect does not limit itself to the particular positive conditioned reflex which undergoes experimental extinction or conditioned inhibition, but in both cases extends to other conditioned reflexes as well, including even those which are of heterogeneous origin. [p. 87]
The final point of resemblance is that in both cases the inhibitory process can be rapidly though temporarily removed, the inhibited reflexes undergoing dis-inhibition on account of external inhibition resulting from alien reflexes to extra stimuli of small intensity. The only point of difference is that in extinctive inhibition it is the positive conditioned stimulus taken by itself which changes its positive significance to an inhibitory one, while in the case of conditioned inhibition the positive conditioned stimulus becomes .involved in a new complex and changes its character in conjunction with an additional stimulus.