An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE VI
Internal inhibition (continued): (c) Delay.
We shall now consider the third type of internal inhibition, which has been termed inhibition of delay. It is obvious from the previous discussion that a considerable choice is allowed in the establishment of conditioned reflexes as regards the time interval between the beginning of the conditioned stimulus and the moment at which it is reinforced by the unconditioned reflex. This interval can be made very short, 1-5 seconds, or even a fraction of a second, provided that the beginning of the conditioned stimulus precedes the moment of application of the unconditioned stimulus. Alternatively, the length of time of the isolated action of the conditioned stimulus can be made comparatively long, extending over several minutes. The reflexes which develop with these two different methods, i.e. with short and with prolonged duration of action of the conditioned stimulus, present great differences with respect to their general properties and their latent periods. These two types of reflex are designated respectively simultaneous -- or to be more precise, almost simultaneous -- reflexes , and delayed reflexes. The duration of the isolated action of the conditioned stimulus is of fundamental importance; since, in the first place, it determines, as we shall see later, the eventual character of every conditioned reflex, and secondly, because it forms the basis of development of that type of inhibition which is the subject of our present lecture.
In all conditioned reflexes in which the interval between the beginning of thc conditioned stimulus and the moment of its reinforcement is short, say 1-5 seconds, the salivary reaction almost immediately follows the beginning of the conditioned stimulus. On the other hand, in reflexes which have been established with a longer interval between the two stimuli the onset of the salivary response is delayed, and this delay is proportional to the length of the interval between the two stimuli and may even extend to several minutes. [p. 89]
Delayed reflexes can be established in various ways. One way is to start by the preliminary establishment of an almost simultaneous reflex -- i.e. one in which the conditioned response appears quickly, say 1-3 seconds after the beginning of the conditioned stimulus -- and to develop the delay gradually. By retarding the moment of the application of the unconditioned stimulus about five seconds each day a corresponding and progressive delay is easily obtained, and we can stop finally at a convenient interval when the required length of delay has been reached.
Another method of establishing a delayed reflex is to pass directly from an almost simultaneous reflex to one with a long delay, leaving out all intermediate stages. As a result of this modification in technique the mode of formation of the delayed conditioned reflex is considerably altered. The reflex, however well established as a simultaneous reflex, disappears at first altogether -- or, to use an expression commonly employed by some of my collaborators, there follows a prolonged period of zeros. Eventually, however, some conditioned secretion of saliva does appear, but not until just before the moment when the unconditioned stimulus is usually applied. On continuing the experiments with the chosen interval of time the secretion progressively increases, and at the same time its commencement shifts further along towards the beginning of the conditioned stimulus and finally settles at a definite intermediate position between the commencement of the conditioned stimulus and its reinforcement.
In both the foregoing methods the experiments start with the establishment of simultaneous reflexes which are then changed either at once or by gradual stages into delayed reflexes. It has been found impracticable, in the great majority of animals, to develop a delayed reflex without first establishing the corresponding simultaneous reflex, since, as will be described in a later lecture, the dogs under these conditions quickly become subjected to drowsiness and sleep so that the experimental formation of conditioned reflexes becomes difficult if not impossible. For this reason the method has received little attention in our investigations.
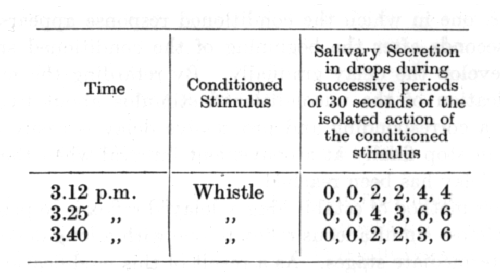
The following example of a delayed reflex is taken from a paper by Dr. Zavadsky who carried out a considerable number of experiments upon internal inhibition of delay. The sound of a whistle is employed as a conditioned stimulus to acid; the isolated action of the whistle is continued during an interval of 3 minutes and is then [p. 90] reinforced by acid, the sound being continued for some time longer so as to overlap the action of the unconditioned stimulus.
The rate of formation of a delayed reflex is subject to great variation. In the first place, the individual character of the animal's nervous system plays an important part. In some dogs the establishment of the reflex is rapid, while in others the beginning of the salivary secretion persistently refuses to separate itself from the beginning of the conditioned stimulus, and the development of delay is very slow. In some animals an indication of a developing delay can be observed in the: course of a single day and after only a few delayed reinforcements; in others there is no indication of the conditioned delay even after a month of persistent work. In the type of dog in which the formation of delay is rapid, it is found that the delay frequently passes into sleep at an early stage of the isolated action of the conditioned stimulus. On this account it is necessary when carrying out a systematic investigation with dogs of this character to restrict the experiments to short-delayed reflexes, which means that we must be content with recording only the small secretion of saliva during the comparatively short time of isolated action of the conditioned stimulus as compared with the larger secretion accompanying a prolonged action of the stimulus.
Another influence affecting the development of delay in the conditioned reflex response is the type of conditioned stimulus used. Tactile and thermal stimulation of the skin and visual stimuli lead to a quicker formation of delay than auditory stimuli, but other things being equal, tactile, thermal and visual stimuli give s smaller total conditioned effect. These facts are illustrated in the following experiments by Dr. Iacovleva:
Preliminary to the development of long-delayed reflexes three short-delayed conditioned alimentary reflexes were established by [p. 91] repeating the isolated action of the stimuli during- 30 seconds, followed by reinforcement. The stimuli corresponding to the three reflexes were the sound of a metronome, a tactile stimulus, and the flash of a lamp.

During the year which intervened between this experiment and the experiment of 24th April, 1925, which is given next, the three reflexes had been used repeatedly in other investigations and had been gradually converted into reflexes of longer delay. The isolated action of the conditioned stimulus had been first prolonged to one minute and then to two minutes, and each reflex had been reinforced an equal number of times.

In the cases of the tactile and visual stimuli the delay is more precise, the reflex response having more perfectly separated itself from the beginning of the conditioned stimulus and being more perfectly related to the time of administration of the unconditioned stimulus.
Another factor which exerts a great influence upon the development of a long-delayed reflex is the amount of practice which has been given to the reflex during the preliminary stage of short delay; [p. 92] a long practised short delay sometimes operates as a persistent obstacle to the development of a longer delay.
Finally, I have evidence that the delay for a stimulus of one and the same character develops at a different rate according as the stimulus is continuous or intermittent. In the former case the development of delay is more rapid.
It can be seen that the reflex response in the case of reflexes with a prolonged delay consists of two phases, an initial phase of inactivity and a subsequent phase of activity, and we must now inquire into the nature of these phases. Does this first and comparatively long period of inactivity mean that the excitatory process is undergoing a progressive summation so that it can evoke an obvious activity when it has reached the necessary intensity? Or is the excitatory process sufficiently strong from the beginning, but unable to produce any secretory effect because it is temporarily overcome by some antagonistic process?
The first possibility must be ruled out straightway on the evidence already given with regard to the formation of delayed reflexes, for seeing that in the case of a short delay the conditioned stimulus can evoke a conditioned reflex with great ease, there seems to be no reason why the same stimulus when more prolonged should require a greater period of summation.
Any suggestion that this first phase of inactivity is caused through fatigue can also easily be eliminated. If, owing to the isolated action of the conditioned stimulus, fatigue did develop, we should expect a gradual diminution of the positive effect of the conditioned stimulus: this, however, is not observed. On the contrary, although the beginning of the secretion gets delayed, nevertheless the secretion increases progressively in amount when the delayed reflex is developed by the first method. With the second method, in which the isolated action of the conditioned stimulus is prolonged from the start, although the positive effect disappears at first altogether, it reappears after a while and then the secretion increases steadily until it settles at a constant maximum value.
There remains now only the supposition that the initial phase of inactivity is due to the excitatory process being temporarily inhibited. That this is the correct interpretation is evidenced by the fact that the existence of an excitatory process in a concealed form during the period of delay can easily be demonstrated: if during the inactive phase of a delayed reflex we act upon the animal by some [p. 93] extra stimulus which has not hitherto been associated in any way with an activity of the salivary glands, we shall immediately elicit a secretion of saliva which is frequently copious and which is always accompanied by the motor reaction peculiar to the conditioned stimulus which was used; in other words, the conditioned reflex becomes revealed throughout the entire duration of the conditioned stimulus in a single positive phase instead of in two phases -- negative and positive. The following are some experiments by Dr. Zavadsky bearing upon this question:
Tactile stimulation of the skin is used as a conditioned stimulus for acid. The conditioned stimulus is allowed to act for a period of 3 minutes and is then reinforced, being still continued so as to overlap the action of the acid. The sound of the metronome which is used in the following experiment has had hitherto no relation to any secretory reflexes and has of itself been unable to evoke any secretion of saliva.

The next example is taken from a further experiment in which a noiselessly rotating object was used as an extra stimulus hitherto neutral with regard to its effect upon the salivary secretion.

[p. 94]
These experiments of Dr. Zavadsky are important as revealing a new and unexpected phenomenon. The established conditioned stimulus had no positive effect by itself during 1 - 1-1/2 minutes, but the neutral stimulus which was added to it for the first time immediately disclosed the regular positive conditioned reflex. It is obvious that we have come across a fresh case of dis-inhibition.
During the last three lectures considerable stress has been laid on the phenomenon of dis-inhibition and many examples have been given. I would plead as my excuse the great importance of an adequate conception of the rôle played by this phenomenon in the physiology of the hemispheres, although of course the study of the lower parts of the nervous system presents many instances of analogous phenomena. So far, however, as the interpretation of the intimate mechanism of dis-inhibition goes we are completely in the dark, and it can only be hoped that accumulation of experimental evidence may at some future date throw light on its nature.
I wish now to call your attention again to the first of the two experiments just described. It can clearly be seen that the stimulus of the metronome when added to the tactile stimulus not only elicited a flow of saliva during the initial phase of inactivity, but also caused a considerable diminution in the salivary secretion during the active phase. While the tactile stimulus applied singly elicited a secretion during the second 1 1/2 minutes ranging from 29-46 drops, the addition of the metronome reduced this to 17 drops. There must, therefore, be a double effect on the part of the extra stimulus -- dis-inhibitory in the initial phase; usually of inactivity; inhibitory in the succeeding phase, usually of activity.
If different extra stimuli producing external inhibition are allowed to act upon delayed conditioned reflexes, various definite and regular modifications are observed in the course followed by the delay. The extra stimuli which were used in this connection for one experimental animal have been arranged in the following groups according to the influence they exerted upon delay:
I. Thermal stimuli at 5"C. and at 440C.; a weak odour of camphor.
II. Thermal stimuli at 0·50C. and at 50"C.
III. Noiselessly rotating objects; the sound of a metronome; tactile stimulation of the skin (the conditioned stimulus to the delayed reflex being in this animal a similar tactile stimulation of [p. 95] a different place on the skin) ; a whistle of moderate strength; the odour of amyl acetate.
IV. Intense odour of camphor; loud whistle; sound of an electric buzzer.
Extra stimuli belonging to the first group did not, in this dog, affect either phase of the delayed reflexes. Extra stimuli belonging to the second group exerted an effect only upon the initial phase of the reflex, causing a salivary secretion. Extra stimuli belonging to the third group disturbed both phases of delay: during the first phase a salivary secretion was produced, and during the second phase the secretion which should normally have been present was much diminished. Extra stimuli belonging to the fourth group exercised little or no influence upon the initial phase of the delayed reflex, but completely suppressed the second phase. It may be added that when all due precautions were taken the experiments proceeded as a rule with striking precision.
The following experiments serve to illustrate the grouping of extra stimuli given above:
A tactile stimulation of the skin is used as a conditioned stimulus to acid in a delayed reflex. The conditioned stimulus acts continuously during three minutes before the administration of acid. The extra stimuli employed are: (1) a thermal cutaneous stimulus of 44°C., (2) a thermal cutaneous stimulus of 0.5°C., (3) an odour of amyl acetate, (4) the sound of an electric buzzer.

[p. 96]

Absolutely identical results are obtained when the extra stimuli are allowed to act, not through the whole of the time of the isolated action of the conditioned stimulus, but only during either the first, inactive phase or the second, active pages. This is illustrated in the following experiments carried out in the same dog and under the same conditions as before.

[p. 97]
The extra stimulus of the whistle, which belongs to the third group of external inhibitors, when acting during the first phase of the delayed reflex brought about a secretion of saliva, but when acting during the second phase it diminished the secretion. A certain diminution of the secretion is sometimes observed during the second phase of the reflex in experiments where the extra stimulus is applied during the first phase. This must be due to the after-effect of the extra stimulus.
It now remains to interpret the empirical grouping of the extra stimuli, and to determine the reason of their different action upon the delayed reflexes. All the experimental evidence at our disposal indicates that the intensity of the effect which they produce depends on their relative physiological strength and the magnitude of the general reactions by which the animal responds to their independent action. The distribution of extra stimuli among the four groups represents a classification according to such physiological strength. In some cases this can be seen by a casual glance at the list itself, where different intensities of the same stimulus appear in different groups.
The difference in the effects of these extra stimuli is also revealed by the motor reaction of the animal (investigatory reflex). With stimuli belonging to the first group there is frequently no motor reaction at all. As we pass on to stimuli belonging to the remaining groups the reactions become more and more vigorous and prolonged. Moreover, the inhibitory effect of all these extra stimuli, which belong, of course, to the group of external inhibitors, is clearly seen in relation to the second (usually active) phase of the delayed reflexes. The inhibitory effect becomes the more pronounced as we ascend from the first to the fourth group until with stimuli belonging to the latter group complete inhibition is obtained. It is thus obvious that the classification represents stimuli of progressively increasing physiological effect upon the organism, those in the first group exerting the least effect.
In cases where an extra stimulus is allowed to act upon a delayed conditioned reflex repeatedly, it is found that its inhibitory effect upon the second (usually active) phase progressively diminishes. This is a further proof that inhibition and dis-inhibition of delayed reflexes by extra stimuli is closely associated with external inhibition. Two experiments may be given which were performed on the dog employed for the last experiment: [p. 98]

It can be seen from these two experiments how the inhibitory effect of the extra stimulus upon the active phase falls gradually with each repetition, the recovery of the delayed reflex being especially regular in the second experiment.
All these experiments, which were carried out on one and the same animal, have been repeated with similar results on many other dogs. The only variation was in the distribution of stimuli among the four groups, which differed slightly for the individual animals. This appears to only reasonable when it is recalled that the intensity of the reactions of different animals in response to extraneous stimuli varies to a considerable and some time to an extreme degree, depending on the individual character of the nervous system and on the previous history of the animal.
Thus we come to the conclusion that variations in the effect of different extra stimuli upon the delayed reflex are determined by differences in the physiological strength of the stimuli. When the strength of the extra stimulus is insignificant the delayed reflex in either of its two phases remains unaffected. When the strength of the extra stimulus is somewhat increased it is only the initial (usually inactive) phase which becomes affected, being now converted into a phase of activity. With a greater strength of the extra stimulus the second (usually active) phase of the delayed reflex also becomes [p. 99] involved, the secretion being considerably diminished in magnitude, so that it becomes equal to or even smaller than the secretion during the dis-inhibited first phase. Finally, with a maximal intensity of the extra stimulus all conditioned activity disappears, and the delayed reflex is represented throughout both phases by a series of zeros.
As a result of the different effects upon the two phases of the reflex brought about through external inhibition, two facts relating to the central nervous activities stand out clearly. The first is that the extraneous stimulus acting on the positive phase of the reflex inhibits, and acting on the negative phase dis-inhibits, in either case, therefore, reversing the nervous process prevailing at the time. The second is that the inhibitory process is more labile and more easily affected than the excitatory process, being influenced by stimuli of much weaker physiological strength.
The following was a chance, but instructive, observation with regard to the second point. I meant to illustrate a series of public lectures on conditioned reflexes by demonstration of experiments. The lectures were given at a place remote from our laboratories so that the dogs had to be conveyed and set down in surroundings which were quite unfamiliar. All the five or six experiments with positive conditioned reflexes, in the first lecture, were carried out successfully. On the other hand, in the second lecture, the experiments with inhibitory reflexes, again five or six in number and attempted on the same animals as before, did not succeed, all the reflexes having undergone dis-inhibition. Thus the very same extra stimulus of the changed environmental conditions had not the slightest disturbing effect upon the positive conditioned reflexes, but exercised a profound influence upon the inhibitory conditioned reflexes, even though the effect of the extra stimulus was now weakened on account of repetition. These facts will be discussed more fully in a further lecture, in connection with other observations bearing on the interrelations between excitation and inhibition.
Since the first phase of a delayed reflex, like experimental extinction and conditioned inhibition, involves an inhibitory process, we should expect to find between all three a close similarity. It has already been noticed in the cases of extinction and conditioned inhibition of a definitely positive conditioned reflex, that the inhibition spreads spontaneously to other conditioned reflexes, and that the degree of this spreading is determined by the relative physiological intensity of the reflexes. It has further been observed that if the [p. 100] reflexes involved secondarily are physiologically weaker than the reflex in which the inhibition was developed primarily, the accompanying secondary inhibition of these other reflexes is complete; but that if the reflexes involved secondarily are the stronger only a partial inhibition is obtained. In other words, the intensity of the primary inhibition is found to be exactly proportional to the intensity of the excitatory process on which it is based. This holds good also for inhibition of delay. An alteration in either direction of the strength of the conditioned stimulus causes a sharp disturbance in the established relation between the inhibitory and excitatory phases of the delay. The two following experiments on this point were conducted on the same animal as was employed in the preceding experiments:
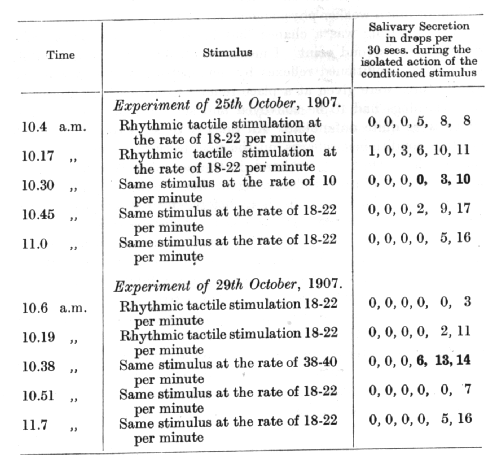
The experiments show a more definite effect in the case of a diminution in the strength of the conditioned stimulus than in the case of an increase in strength, though both effects are evident. [p. 101]
Just as with extinction and conditioned inhibition, the intensity of the inhibition in delay also depends upon the strength of the unconditioned stimulus. This can easily be shown with conditioned alimentary stimuli, when the delayed reflex is tested in the dog after it had been fed at the usual time and again after a certain period of fast. The following experiment illustrates this point. A whistle of moderate strength is used as a conditioned alimentary stimulus in a conditioned reflex delayed by three minutes.

Experiments of this kind show that when the physiological significance of the conditioned stimulus is increased through deprivation of food, the inactive or inhibitory phase of the delay almost entirely disappears.
Summation of intensity of the inhibitory after-effect can be observed with inhibition of delay exactly as with extinction and conditioned inhibition. This is clearly evidenced by an experiment such as the following:
A tactile stimulation of the skin serves as a conditioned stimulus in a delayed reflex, the isolated action of this stimulus being [p. 102] continued during three minutes before the application of the unconditioned.

[p. 103]
The tactile stimulus which normally produced inhibition during the first 1 1/2 minutes of its isolated action, when abbreviated so as not to act during the second 1 1/2 minutes associated with the excitatory phase of the reflex, led to such a strengthening of the inhibition that on its subsequent application for the full three minutes the positive phase of the reflex was either greatly reduced (11.21 a.m.) or else abolished altogether (12 noon).
It remains now to seek an interpretation of the fact that in the case of a delayed reflex, the same stimulus has at first an inhibitory, and later an excitatory, effect. What factor determines these two distinct properties of one and the same stimulus acting under apparently identical conditions? We shall find no difficulty in correlating this phenomenon with the experimental evidence already considered in these lectures. In the third and fifth lectures a number of agencies were discussed which can be given either excitatory or inhibitory conditioned properties. In particular the factor of duration of time was shown to act as a real physiological stimulus, and experiments were described in which definite time intervals appeared as effective stimuli. I should like especially to recall to your memory the experiment in which the compound stimulus consisted of an external stimulus related to a definite moment of time (p. 41). In this experiment the external stimulus was without any effect until applied in the neighbourhood of the particular moment after the previous administration of acid, but as this particular moment was approached the secretory effect made its appearance and gradually and precisely increased to a maximum. This is exactly the case with delayed reflexes also. In the experiments which have just been described in the present lecture the unconditioned stimulus was added to the external stimulus only at the expiration of 3 minutes; in other words, the external stimulus itself plus its duration for three minutes together constitute the actual compound stimulus which was immediately reinforced, and it was this particular combination and not the nominal conditioned stimulus which acquired conditioned properties. The same nominal stimulus at any time previously to the end of the third minute acted as a component of a different stimulatory compound which remained unreinforced by the unconditioned reflex and therefore became inhibitory, exactly as would any other stimulus if it were not reinforced. The same phenomenon has already been demonstrated in the case of conditioned inhibition, and illustrations even more [p. 104] striking will be given in the next lecture, which is to deal with the fourth type of internal inhibition.
In the case of delayed reflexes the significance of the duration of a stimulus can be observed in a very concrete and simple manner. When the external stimulus selected for the formation of a conditioned reflex is applied during a given interval of time, at each successive moment the stimulus forms part of a definite and distinct stimulatory compound. It is well known how soon we get accustomed to stimuli of smell, sound or illumination. This, of course, means that the nerve cells which are being excited pass through a series of successive physiological changes. In accordance with this it is obvious that if a definite unconditioned reflex is repeatedly evoked coincidently [sic] with any one particular physiological state of the cerebral cells, it is this definite state and no other that acquires a definite conditioned significance. In the next lecture it will be shown to how great an extent the discrimination of different intensities of one and the same stimulus can proceed, and how the stimulus evokes at one particular intensity a positive and at another intensity an inhibitory conditioned reflex. We are thus fortunately provided with a great deal of perfectly good experimental evidence which throws light on the phenomenon of internal inhibition involved in delay.
The possibility of the development of a delay has always to be reckoned with in studying conditioned reflexes. We know that in order to determine the intensity of excitation in a simultaneous or a short-delayed conditioned reflex in its numerous and subtle variations under different conditions, it is necessary to apply the appropriate conditioned stimulus singly for a longer or a shorter period of time, combining its action with that of the unconditioned stimulus at the end of this interval. Only in this manner can we get a measurable reflex response. This procedure, even though rarely performed, tends to the development of a more or less prolonged delay, so that along with the excitatory process there originates also an inhibitory one. The investigation thus naturally becomes complicated, for it is now necessary to deal with two simultaneous and antagonistic processes. For example, it is not easy by our usual methods to obtain the true latent period of conditioned reflexes, since what has often been referred to in our description of experiments as the "latent period" represented in actual fact a delay -- i.e. the interposition of an inhibitory period, which could appropriately [p. 105] be termed the "preliminary inhibitory period," but which is certainly not the true latent period of the reflex.
In order to determine the true latent period of conditioned reflexes it is necessary to use reflexes which are as nearly as possible simultaneous, the unconditioned stimulus succeeding rapidly on the conditioned after an interval of a fraction of a second. It is only by using such reflexes, and then chiefly their motor components, that convincing experimental evidence can be obtained showing that the length of the true latent period of a conditioned reflex is really of the same order as is the true latent period of reflexes in the lower centres of the brain and spinal cord. Such determinations have only recently been introduced into our investigations, as we did not consider the determination of the true latent period of conditioned reflexes to be of fundamental importance for establishing their nature as reflex. The regularity and infallibility with which these reflexes could be evoked constituted in our opinion the main evidence for their reflex nature, and in this connection differences in the period of latency did not seem of much importance since the latent period of lower reflexes also is known to be subject to fairly wide variations depending on the complexity of the central paths and connections involved in the reflex. It may justly be admitted, however, and without prejudice to our conception of the reflex nature of conditioned reflexes, that the nervous connections are of greater complexity in those reflexes which involve the cerebral cortex.
The elucidation of many other problems concerning conditioned reflexes, such, for instance, as that of the actual course of the excitatory process from the moment of its origination, is also rendered difficult by interference from delay. In this respect individual differences in the character of the nervous system may be very helpful. For example, it was shown above that in some animals the delay is developed with difficulty, and that the excitatory process is only slightly, or often not at all, disturbed. The experimenter, in order to diminish the influence of delay, can also take the precaution to abbreviate the isolated action of the conditioned stimulus to the minimum compatible with obtaining a secretion of sufficient magnitude to allow of comparison in different variations of the experiment. On the other hand, there are some problems in the elucidation of which the initial process of inhibition may be turned to advantage, as will be shown in the next lecture. It can clearly be seen how, when inquiring into the properties of conditioned reflexes, the [p. 106] experimenter has to adapt his methods to the character of the animal at his disposal.
The experimental evidence with which we have been dealing in the last three lectures demonstrates the enormous biological importance of internal inhibition of conditioned reflexes. It is by means of internal inhibition that the signalizing activity of the hemispheres is constantly corrected and perfected. To sum up:
If over a given time a signalling, i.e. a conditioned, stimulus is repeatedly presented without the accompaniment of the unconditioned stimulus, then the conditioned stimulus becomes meaningless to the organism as calling for an unnecessary expenditure of energy, and the stimulus loses, though generally for only a short time, its physiological significance (experimental extinction).
In exactly the same manner, if the conditioned stimulus is repeatedly applied together with another extraneous stimulus and in the combination is never followed by the unconditioned stimulus, the conditioned stimulus loses its positive excitatory conditioned effect, but only in that particular combination and not when applied singly (conditioned inhibition).
Finally, if a regular interval of sufficient duration is established between the commencement of a conditioned stimulus and its reinforcement by the unconditioned stimulus, the former becomes ineffective during the first part of its isolated action; during the second part of its action a positive excitatory effect appears, and this increases progressively in intensity as the moment approaches when the unconditioned stimulus has customarily been applied (inhibition of delay).
In the above manner a continuous and most exact adaptation of the organism to its environment is effected, revealing a most delicate adjustment in the antagonistic nervous processes of the higher animals.
As has been clearly demonstrated in the last three lectures the phenomena, of extinction, conditioned inhibition and delay all represent the formation of inhibitory conditioned reflexes. Inhibitory conditioned reflexes can, however, also be obtained by a totally different procedure. If an inhibitory stimulus is applied simultaneously and repeatedly for short periods of time together with some neutral stimulus the latter also develops an inhibitory function of its own.
This subject has been specially examined by Dr. Volborth, who started by rendering the extraneous stimuli absolutely neutral, [p. 107] presenting them repeatedly to the animal until they ceased to produce any inhibition of the positive conditioned reflexes or any dis-inhibition of the inhibitory reflexes. He then repeatedly allowed these neutral stimuli to act during short intervals of time either with conditioned reflexes which had just been experimentally extinguished or with the inhibitory combination in a conditioned inhibition. After repeating this procedure several times he tested the action of these hitherto neutral stimuli upon positive conditioned reflexes, and in this manner successfully demonstrated that they had acquired definite inhibitory properties.
Since in order to become sure of the results these tests had to be repeated several times, Dr. Volborth in one group of experiments accompanied only every alternate test by the unconditioned stimulus. This strict alternation of reinforcement with non-reinforcement was adopted in order to prevent the combination of the positive stimulus with the new and hitherto neutral stimulus from acquiring a predominance of positive or negative properties in itself. But most convincing of all was another form of the experiment in which the test was always accompanied by the unconditioned stimulus. In spite of repeated tests of this kind the recently acquired inhibitory properties of the hitherto neutral stimuli were still prominently exhibited. The following are some of Dr. Volborth's experiments, in which a "natural" alimentary stimulus was employed.
The alimentary conditioned stimulus after extinction was repeatedly applied in conjunction with the sound of a metronome which had previously been rendered entirely neutral. After this procedure had been continued for some time a combination of the action of the metronome with a positive and non-extinguished alimentary reflex was occasionally tested, and this combination was always reinforced by the unconditioned reflex.

[p. 108]
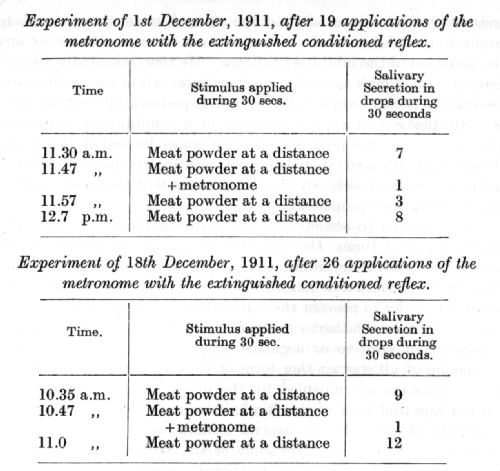
It is obvious that the sound of the metronome which was formerly entirely neutral has acquired inhibitory properties as a result of repeated applications with the extinguished conditioned reflex.
This new inhibitory stimulus of the second order proves itself in every respect similar in properties to those inhibitory stimuli which have been considered already in connection with experimental extinction, conditioned inhibition and delay. For example, a new inhibitory stimulus of the second order which is developed with the help of one definite conditioned reflex can exert an inhibitory effect upon other conditioned reflexes as well. This is shown in the following experiment of 15th March, 1911, in which a different dog is employed.
The sound of a metronome was made always to coincide with an extinguished "natural" conditioned alimentary reflex; subsequently the effect of the metronome was tested upon an artificial alimentary conditioned reflex to odour of camphor. [p. 109]

It can be seen also in the above experiment that the effect of the conditioned inhibitory stimulus of the second order is not limited to the time of its actual administration, but reveals itself also in a definite inhibitory after-effect which is subject to summation. Furthermore, conditioned inhibitory stimuli of the second order are, equally with the primary ones, subject to dis-inhibition by agencies belonging to the group of external inhibitors. We thus come to the following conclusion: when perfectly neutral stimuli fall upon the hemispheres at a time when there prevails a state of inhibition they acquire an inhibitory function of their own, so that when they act subsequently upon any region of the brain which is in a state of excitation they produce inhibition.
A point to which further reference will be made should be mentioned here, namely, that every extraneous stimulus which falls upon the hemispheres and remains without any further consequence to the animal, if repeated causes the spontaneous development of a cortical inhibition. Therefore, in the type of experiments just described, it is always necessary to ascertain the extent to which the acquisition of inhibitory properties by hitherto neutral stimuli depends on their simultaneous application with inhibitory stimuli, and to what extent these inhibitory properties have developed independently.