
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE VIII
The analysing and synthesizing activity of the cerebral hemispheres (continued): (c) Examples of the analysis of stimuli. (d) Synthesis and analysis of compound simultaneous stimuli. (e) Synthesis and analysis of compound successive stimuli.
It was shown in the preceding lecture that the animal at first generalizes any definite individual stimulus of the outer world, but that with repetition the stimulus becomes more and more specialized as a result of the development of an inhibitory process (differentiation). In this final form conditioned reflexes provide s reliable method for an experimental study of the scope and limits of the activity of the different cortical analysers. Our knowledge of the different analysers in the dog has recently been considerably advanced through the study of conditioned reflexes, -- a fact which affords a, striking example of the practical utility of this method of research. It should, moreover, be mentioned with regard to these particular experiments, that while considerable difficulties were sometimes encountered in the course of the work, these did not arise in the physiological part of the technique, but derived from instrumental limitations, since in many cases it has been exceedingly difficult to obtain or construct suitable physical apparatus. The main requirement is for instruments which are capable of producing a perfectly isolated and unvarying elementary stimulus of a definite degree of intensity. This, however, is very often a practical impossibility. For example, it is exceedingly difficult to find an apparatus for tactile stimulation of the skin which will not produce some slight sound during its application. It is also not easy to obtain an alteration in the pitch of a tone without simultaneously affecting its strength. Indeed, it seems to me that future experimentation upon the analysers of animals will exhibit an interesting competition between the delicacy of the nervous analyser and the skill of the instrument maker.
We shall turn now to a consideration of the experiments at our disposal, taking first those dealing with the visual analyser of the dog. In respect of discrimination of luminosity this analyser was [p. 132] found to be greatly superior to that of man. Thus, for example, a conditioned reflex was established in a dog to the presentation of a black screen perfectly uniform in shading, without any traces of graining or spots. A white screen of identical shape and size, and also of uniform luminosity, was differentiated from the black screen by the usual method of contrast. The experimenter was provided with a number of screens of different shades (50 numbers of Zimmermann's collection) ranging from white through different shades of grey to black. After the differentiation of white had been firmly established the same method of contrast was used to obtain finer degrees of differentiation of grey screens approximating more and more nearly to black. It was found in this way that the visual analyser of the dog was capable of distinguishing between the neighbouring shades Nos. 49 and 50, while to the human eye there appeared not the slightest difference between them, whether they were examined successively at different intervals of time or simultaneously. This was also true for several other shades separated further from one another on the scale, which could not be discriminated by the human eye but which the dog differentiated perfectly. The following experiment shows an evident, though not absolute, differentiation of the screens Nos. 49 and 50 [experiments by Dr. Frolov]:
It thus becomes evident that as regards the analysis of intensity of illumination the visual analyser of the dog is so highly developed that we were unable to determine the limit to which this activity extends .
In the case of analysis of various colours the results obtained were quite different. Dr. Orbeli in s first series of experiments was unable to detect any differentiation of colours on the part of his dogs. In a second series of experiments, however, positive results were obtained in one dog, but only with great difficulty, and even in this case the experiments were still open to criticism. The results obtained [p. 133] by other investigators, both Russian and foreign, lead to the conclusion that colour vision in dogs, if present, is only of a very rudimentary form, and that in most dogs it cannot be detected at all.
Dr. Orbeli studied also the differentiation of figures. Examples of figures for which an absolute differentiation was obtained, are given in Fig. 6.
Experiments with regard to differentiation of shapes were continued by Dr. Shenger-Krestovnikova . An alimentary conditioned reflex was established in a dog to s luminous circle which was projected on to a screen placed in front of the dog. After the reflex had attained a constant strength the animal was made to differentiate from the circle a number of ellipses of equal surface and luminosity. In the first of the ellipses the ratio of the semi-axes was 2: 1, and differentiation was established with ease. This was followed up by a series of ellipses which gradually approximated to the circle in shape, and so required a finer and finer differentiation. The ellipse with ratio of the semi-axes 9:8 proved to be the limit at which differentiation just failed. Some indication of differentiation appeared at first, but on repetition it gradually disappeared, and with it disappeared also all the previously established coarser differentiations. To renew these it was necessary to work up carefully from the very beginning, starting with the first ellipse with a ratio of the semi-axes 2:1. When all the coarser discriminations had again been obtained, the ellipse with the ratio 9:8 was tried once more. Its first application showed a complete discrimination giving a zero secretion of saliva. Further tests, however, led to the same results as before. Not only was it impossible to obtain the differentiation again (if the first trial can be regarded as a real differentiation at all), but all the earlier, coarser differentiations disappeared as well. In this phenomenon we have a clear reproduction of the case referred to at the end of the preceding lecture. When the stage of minute differences between stimuli is reached, analysis of itself appears still feasible, but the relations existing between the excitatory and the inhibitory processes seem to present an insurmountable obstacle to its continued and permanent utilization by the animal for an appropriate responsive activity.
The differentiation of direction of motion of figures and points was also investigated in our laboratories, but the limit of discrimination was not determined in these cases.
The analysing activity of the acoustic apparatus in the dog was [p. 134]
 [p. 135]
[p. 135]
investigated in especial detail and in various directions. In the first place we shall consider the analysis of different intensities of the same sound. It was found that any definite degree of intensity of a sound could easily be made into a stable conditioned stimulus and could be differentiated from slightly higher or slightly lower intensities of the same sound [experiments of Dr. Tichomirov]. A tone of 1740 d.v. was sounded by an organ pipe into which air was blown at a constant pressure of 3.6-3.8 cms. of water by means of a spirometer. The organ-pipe was fitted in the centre of a wooden board covered by a thick layer of cotton wool. Above this board and over the pipe was suspended a wooden box, open below and also coated inside with cotton wool. By raising or lowering this box to different heights over the pipe definite dampings of the sound were obtained. The limit of differentiation to the intensities of a given sound could now be determined for the dog and compared roughly with that of human beings. Thus it was found that an intensity very closely approaching the one employed as a positive conditioned stimulus could be differentiated by the dog with an absolute precision even when a pause of 17 hours was made between the two stimuli. The experimenter found himself able to detect a difference between these two intensities of the sound only when they succeeded each other immediately. The following is an example taken from these experiments:

In the continuation of these experiments the intensity of the inhibitory tone was brought still nearer to the intensity used for the positive conditioned stimulus, and an absolute differentiation was obtained even after a pause of three hours between the stimuli. Unfortunately these experiments were conducted in our old laboratory where the effect of the inhibitory stimulus was easily disturbed, and it must be left to the future to repeat these experiments under more perfect conditions in our new laboratory. [p. 136]
A great number of experiments have also been carried out on the differentiation of pitch. To produce tones of different pitch wind instruments were chiefly employed, and the limit to which we were able to carry our tests was an interval of one-eighth of a tone (800 and 812 d.v.). This interval, as has already been shown in the previous lectures (p. 125), was differentiated with perfect accuracy by the acoustic analyser of the dog. It was not possible to carry our determination of the physiological limit of differentiation of pitch any further, since we could not be sure that our apparatus would accurately reproduce smaller intervals than one-eighth. The repetition of these experiments with the use of pure tones, produced by telephones with resonators, showed that this differentiation could still be obtained as easily as with the richer tones of wind instruments [experiments of Drs. Anrep and Manuilov].
The upper limit of the auditory range of the dog was demonstrated by Dr. Bourmakin with the use of a Galton's whistle, and by Dr. Andréev with the use of an apparatus producing pure tones, to be very much higher than in man. Conditioned reflexes were successfully established to tones of such high pitch as to he quite inaudible to man. It was indeed interesting to observe how sharply and precisely the dog reacted to sounds which were non-existent to the human ear.
The differentiation of timbre and the differentiation of direction of sounds were also submitted to investigation, but no exact limits of differentiation were determined in the cases of these special qualities.
While discussing the functional capacity of the acoustic analyser we may refer to those experiments in which differentiation was based not upon the differences in the properties of the sounds but upon differences in the rhythm of successive applications of one and the same single sound. The sound in this case was produced by a metronome beating regularly, but at different rates in the various experiments. Differentiations of this type were quite easily obtained, but the main interest of these investigations lay in the determination of the limit of differentiation, which for the dog was found to be far more subtle than the discrimination which could be recognized in man. The dog was capable of a very precise differentiation between such rates as 100 and 96 beats per minute even when applied at a very long interval after one another (i.e. a discrimination of 0.024 second).
A few experiments have been conducted with the analysers for tactile and thermal cutaneous stimuli. Differentiation could easily [p. 137] be established for the site of stimulation, and was found to be very precise, but no determination was made as to the actual limit.
Differentiations were obtained also for different types of tactile stimulation, namely pressure by smooth and rough surfaces, pressure by blunt points arranged in various patterns, and by scratchings in various directions with a small brush. Furthermore, differentiations were also obtained for different degrees of temperature.
The most perfect analyser in the dog is the chemical analyser of smell, but our information concerning it is scanty, mainly on account of instrumental difficulties. It has been exceedingly difficult, if not impossible up to the present, to obtain the same accuracy in graduation of olfactory stimuli as of any other stimuli. It is impossible also to limit the action of olfactory stimuli to any exact length of time. Furthermore, we do not know of any subjective or objective criterion by which small variations in intensity of odours can be determined. On this account only a small number of experiments could be conducted. Differentiations were obtained for various odours -- camphor, vanillin and many others. Some of these were made into positive conditioned alimentary stimuli or positive conditioned stimuli for acid, while others were given the corresponding inhibitory properties. Experiments upon differentiation were also conducted with mixtures of odours, into which some new odour could be introduced.
Lastly, some experimental data have been obtained concerning the chemical analyser of taste. In this case matters were more complicated, because the unconditioned stimuli which were usually employed in the experiments (food and rejectable substances), both act on this analyser. In order to study the analyser of taste in a manner similar to that employed for other analysers, it would be necessary to employ an unconditioned reflex belonging to some other analyser and to use various taste stimuli for establishing the corresponding positive and negative conditioned reflexes. Experiments of this kind have not been performed. We adopted, however, another method in which a number of conditioned reflexes were established each associated with a different nutritive or rejectable substance (meat powder, bread crumbs, sugar, cheese, acid, soda, etc.), and their interaction and mutual inhibition were then observed. The following is an example of such an experiment by Dr. Egorov:
A tactile stimulation of the skin presented a conditioned stimulus to the consumption of a mixture of meat powder with bread crumbs; [p. 138] a rotating object was used as a conditioned stimulus to the consumption of Dutch cheese.

Since the conditioned stimulus of the rotating object (3.50 p.m.) was reinforced by administration of cheese, the above form of the experiment does not tell us whether the subsequent diminution in the conditioned reflex to the tactile stimulus (i.e. the conditioned stimulus for meat and bread) was due to the conditioned stimulus of the rotating object or to its reinforcing agent (i.e. cheese). To determine this point similar experiments were conducted with the use of the conditioned stimuli applied without reinforcement. The following is an example of such an experiment.
Tactile stimulation of the skin usually resulted in this dog in a conditioned secretion varying from 5 to 6 drops of saliva during 30 seconds. The stimulus of the rotating object which is given prior to the application of the tactile stimulus remains unreinforced.
 [p. 139]
[p. 139]
The very considerable and protracted diminution in the reflex to the tactile stimulus shows that it is the analysing activity of the cerebral part of the chemical analyser of taste which is involved in this case, since the conditioned stimulus to cheese was in itself sufficient without reinforcement to bring about a profound diminution in the reflex to the tactile stimulus.
Another set of experiments was performed [by Dr. Savich] in which "natural" conditioned reflexes to the appearance of meat powder and to the appearance of granulated sugar were employed. The dog's usual diet outside the experiment consisted of oatmeal porridge with meat and bread. Later on meat and bread were excluded from the meal, but a large amount of sugar was added to the porridge. After this diet had been maintained for some time the conditioned reflex to the appearance of meat powder was found to be considerably increased, while the reflex to the appearance of sugar had disappeared almost entirely.
Similar experiments, but with greater precision of detail, were carried out by Dr. Hasen with conditioned reflexes to rejectable substances. He took advantage of two facts which had previously been noticed in reflexes to acid, viz., that in most of the cases the magnitude both of conditioned and of unconditioned reflexes gradually rises towards the end of an experimental day, and that the magnitude of the reflexes also rises, up to a certain maximum, in the course of a series of experiments conducted on successive days. Dr. Hasen modified his experiments in the following manner. After the first application of the conditioned stimulus, which was reinforced, acid was introduced several times in succession without the conditioned stimulus, and finally the conditioned stimulus was again applied. The effect of the conditioned stimulus on this last application was found always to be increased as compared with the effect on its first application. At a later stage in the experiments, which were performed on two dogs, the daily routine was interrupted by three intervals, of five days each in one series of experiments, and of three days each in another; During the first interval the one dog received in the form of an enema, and the other dog received through a stomach tube, a considerable amount of dilute acid. During the second interval a solution of sodium carbonate was given in a similar manner, and during the thud interval the dogs received no injections at all. The conditioned reflexes and the unconditioned reflexes were tested after each of these intervals. It was found that after the [p. 140] interval in which acid was introduced the magnitude of the reflexes remained the same or was but slightly diminished as compared with the experiments preceding the pause; after the interval in which nothing was administered the magnitude of the reflexes fell considerably; and after introduction of soda the reflexes diminished still more. In the course of daily experiments a definite amount of acid introduced into the mouth of one of these dogs produced from the submaxillary gland on an average a salivary secretion of 5.1 c.c., while the conditioned stimulus elicited a secretion of 4 drops during 30 seconds. After the interval during which nothing was administered the figures were 3.8 c.c. and 2 drops respectively; after the interval in which sodium carbonate was introduced 3.7 c.c. and a secretion of zero were respectively produced; finally, after the interval in which acid was administered, the secretions obtained amounted to 4.5 c.c. and 3 drops respectively. Thus it is evident that the variations in the chemical composition of the blood which resulted from excess of acid or alkali were differentiated by the cerebral termination of the chemical analyser, being manifested by an increase or a decrease in the excitability of the central part of the analyser. When the animal absorbed an excess of acid the excitability of the "acid" part of the chemical analyser increased, with the result that on encountering acid from the outer world the organism responded by more vigorous motor and secretory reflexes directed towards the exclusion of the further introduction of acid. The same, of course, takes place in the case of nutritive substances, a corresponding increase or decrease being observed in the positive or negative reactions to different substances or different quantities of them. It is thus seen that the chemical analyser of taste in its central part forms a connecting link between the internal and external media of the organism, and by regulating their relations one to the other secures a certain constancy of the internal medium.
The experiments which have just been described, and which, unfortunately, were not subsequently repeated, belong to the earlier period of our work, when the novelty and complexity of the subject afforded numerous sources of error. Nevertheless, even so, they show definitely that the study of the chemical analyser of taste, though complex, can be conducted successfully by the use of conditioned reflexes. [p. 141]
With regard to the synthesizing activity of the nervous system, as compared with its analysing activity, little is known up to the present. It would, indeed, be futile for us to attempt to discuss the nature of its intimate mechanism : it can only be suggested that, in the future, synthesizing activity will be referred to the physico-chemical properties of synaptic membranes or anastomosing neuro-fibrils. Our immediate task must consist in accumulating experimental material concerning the synthesizing activity.
In addition to the formation of the conditioned reflex itself -- which is, of course, primarily an expression of the synthesizing activity, and which has constantly formed the starting-point of our investigations-we have also examined the properties of compound conditioned stimuli. Compound stimuli were used with either simultaneous or successive action of their component parts.
In the case of compound simultaneous stimuli the following important relations have been observed :
When the stimuli making up the compound act upon different analysers, the effect of one of them when tested singly was found very commonly to overshadow the effect of the others almost completely, and this independently of the number of reinforcements of the compound stimulus. For example, a tactile component of a stimulatory compound was usually found to obscure a thermal component, an auditory component to obscure a visual component, and so on. Thus, in the following experiment by Dr. Palladin a conditioned reflex to acid was established to a simultaneous application of a thermal stimulus of 0°C. and a tactile stimulation of the skin. Tests were made both of the compound stimulus and of its individual components applied singly.

Another example may be taken from an experiment by Dr. Zeliony. A conditioned alimentary reflex was established to the simultaneous application of the tone of a pneumatic tuning-fork, which was considerably damped by being placed within a wooden [p. 142] box coated with wool, and of a visual stimulus of three electric lamps placed in front of the dog in the slightly shaded room.
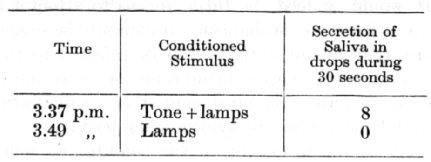
In the above experiments the action of the thermal and visual components by themselves was ineffective, being completely overshadowed by the other respective components. It is obvious, however, that the ineffective components in the stimulatory compounds could easily be made to acquire powerful conditioned properties by independent reinforcement outside the combination. The true interpretation of the phenomenon which has just been described is revealed by experiments in which both components of a stimulus belong to one and the same analyser. For example, in one experiment there were used as components in a stimulatory compound two different tones, which appeared to the human ear to be of equal intensity. When the conditioned reflex to the compound because fully established, the tones sounded separately were found to produce an equal effect. In another experiment, a conditioned reflex was formed to a compound in which the two individual tones were of very different intensities. The effect of the tone of weaker intensity when tested singly was now very small or absent altogether. Such a case is illustrated in the following experiment by Dr. Zeliony. An alimentary reflex was established in a dog to a compound stimulus made up of the sound of a whistle and the sound of the tone d sharp of a pneumatic tuning-fork. Both these sounds appeared to the human ear to be of equal intensity, and both when tested separately elicited a secretion of 19 drops of saliva during one minute. In addition to this, another compound stimulus was established, made up of the same sound of the whistle plus the tone a of a tuning-fork of weaker intensity. When tested separately the whistle in this case elicited a secretion of seven drops of saliva during thirty seconds, and the tone only one drop.
It is evident from the above experiments that the obscuring of one stimulus by another belonging to the same analyser is determined by differences in their strength, and it is natural to assume [p. 143] that this explanation can be applied also to compound stimuli, the components of which belong to different analysers. On this assumption tactile cutaneous stimuli in our experiments should be regarded as being relatively stronger than thermal cutaneous stimuli, and auditory stimuli should be regarded as being relatively stronger than visual stimuli. The natural deductions from such an assumption are far-reaching, and it will be necessary at some future date to test its validity by the use of stimuli compounded from different analysers and varying as much as possible in their intensities, very weak auditory stimulus being combined with a very strong visual one, and so on.
The phenomenon in which one stimulus is obscured by another in a simultaneous stimulatory compound, when the two stimuli belong to different analysers, presents several interesting features. The effect of the compound stimulus is found nearly always to be equal to that of the stronger component used singly, the weaker stimulus appearing therefore to be completely overshadowed by the stronger one. If, however, the stronger stimulus is even at long intervals of time, repeated singly without reinforcement by the unconditioned reflex, while the compound stimulus is constantly reinforced, the stronger stimulus by itself becomes completely ineffective, whereas in the stimulatory compound there is no diminution in its effect. It is evident, therefore, that although the effect of the weaker stimulus when tested singly is invisible, it nevertheless plays an important part in the stimulatory compound [experiments by Dr. Palladin].
Another feature of interest has been already described in the fourth lecture [experiments of Dr. Perelzweig, page 56]. If the weak component, which may be even quite ineffective when applied alone, is repeated at short intervals without reinforcement -- i.e. is extinguished below zero -- then both the compound and the stronger component undergo secondary extinction. In this experiment, therefore, the component which is normally of itself apparently ineffective becomes temporarily transformed by the process of experimental extinction into a strong inhibitory stimulus.
The following was observed in a single, but so far as the experimental conditions were concerned a perfect, experiment. When two stimuli belonging to different analysers were first separately made into conditioned stimuli, and only afterwards applied simultaneously to form a compound stimulus, which was repeatedly [p. 144] reinforced, the overshadowing of the one component by the other did not occur. From this it may be concluded that in the usual case where two hitherto neutral stimuli are used to form a compound stimulus, the stronger stimulus at once prevents the weaker from forming a corresponding connection with the centre for the unconditioned reflex. If, however, this connection has been established already, it is not disturbed during the subsequent establishment of the reflex to the compound stimulus. The mechanism on which the predominance of one component of a stimulatory compound over another depends is most probably a form of inhibition. This matter will be examined in detail in a subsequent lecture.
The cases mentioned above show that a definite interaction takes place between different cells of the cortex, resulting in a fusion or synthesis of their physiological activities on simultaneous excitation. In the case of a compound simultaneous stimulus made up of components of unequal strength belonging to the same analyser this synthesis is not so obvious. However, it comes out very clearly that even in these cases there is no summation of the individual reflex effect of each single component, the effect of the stronger component applied singly being equal to that of the compound stimulus.
The phenomenon of synthesis of stimuli belonging to the same analyser is much more evident in a modification of the experimental conditions which was first used in Dr. Zeliony's experiments, then again by Drs. Manuilov and Krylov, and has since been widely practised. It was noticed that if a conditioned reflex to a compound stimulus was established as described above, it was easy to maintain it in full strength and at; the same time to convert its individual components, which gave a positive effect when tested singly, into negative or inhibitory stimuli. This result is obtained by constant reinforcement of the compound stimulus, while its components, on the frequent occasions when they are applied singly, remain without reinforcement. The experiment can be made with equal success in the reverse direction, making the stimulatory compound into a negative or inhibitory stimulus, while its components applied singly maintain their positive effect. We leave the discussion of this phenomenon to Lecture XVI, and shall pass on to consider the second type of stimulatory compounds, namely, compound successive stimuli. [p. 145]
In this second type of stimulus, in which the component stimuli are applied not simultaneously but in succession, the synthesizing function of the nervous system is still more obvious. The compound stimuli employed were of many different kinds. In some cases the compound was made up of successive repetitions of one and the same stimulus. For example, a definite tone was repeated three times for one second with an interval of two seconds between the first and second applications, and an interval of one second between the second and third; this rhythm was repeated after a pause of five seconds, and was now accompanied by the unconditioned stimulus. In other experiments the stimulatory compound was made up of three or four different stimuli all belonging to one analyser; the stimuli were made to succeed one another in a definite order, being each of equal duration, and with equal pauses between them. There were used, for example, in one case the four tones C, D, E, F of one octave; and in another case the four stimuli were made up of a noise, two different tones and the sound of a bell. Finally, in other experiments there was employed a stimulatory compound composed of three or four stimuli belonging to different analysers, each stimulus being of equal duration, the pauses between them being also equal. Conditioned reflexes were readily obtained to all these different compound stimuli, and after a certain amount of practice of the reflexes all the individual components when tested singly were found to exhibit a positive conditioned effect, which varied. In magnitude according to the quality and relative strength of the individual stimuli.
The next step was the introduction of different modifications of these compound stimuli. In the first case the order of the two pauses between the repetitions of the tone was reversed, the longer pause being now made between the second and third applications instead of between the first and second. In the remaining cases the order in which the different stimuli were applied was changed, either completely by reversing it, or in the case of the compound consisting of four component stimuli by reversing the order of the two middle ones. These modified compounds were repeatedly applied without reinforcement, but when the stimuli were applied in their original order the compound was always reinforced, with the result that ultimately the original compounds became differentiated from their modifications, which latter finally lost their positive conditioned effect and acquired an inhibitory one [experiments [p. 146] by Drs. Babkin, Stroganov, Grigorovich, Ivanov-Smolensky and Eurman].
The following is an example of the different permutations of four tones (1, 2, 3, 4) which were successfully differentiated by the dog in experiments by Dr. Babkin, the sequence 1234 being the positive stimulus and the sequence 4321 the first stimulus to be differentiated; this was followed by development of an absolute differentiation of all the remaining sequences. The vibration frequencies of the four tones employed were 290, 325, 370 and 413 d.v. respectively.

In the following experiment [by Dr. Eurman] the positive alimentary conditioned stimulus was made up of the flash of an electric lamp (L), a tactile cutaneous stimulation (C), and a sound of bubbling water (S),applied in that order, namely, L-C-S. The inhibitory compound was made up in the reverse order, namely, S-C-L.

The following is an experiment by Dr. Ivanov-Smolensky. The positive conditioned alimentary stimulus was made up of a hissing sound (H), a high tone (hT), a low tone (lT), and the sound of a buzzer (B); applied in that order, namely H-hT-lT-B. The inhibitory stimulus was made up with the order of the two middle components reversed, namely H-lT-hT-B. [p. 147]
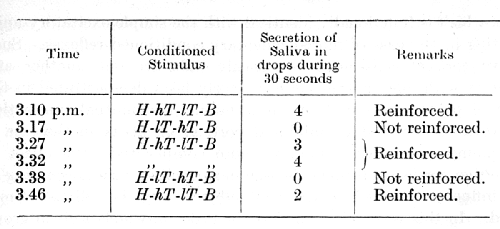
The formation of these inhibitory reflexes usually required great deal of time: although a relative differentiation could sometimes be observed quite early, absolute differentiation was obtained in extreme cases only after more than one hundred repetitions without reinforcement. Indeed, in order to obtain a complete differentiation it was sometimes necessary to proceed by gradual stages, commencing with differentiation of comparatively simple compound stimuli. It was especially difficult to obtain a differentiation of the stimulatory compound of sound, low tone, high tone and buzzer from the compound in which the tones were reversed. All the differentiations, and especially those exceptionally difficult of formation, proved very unstable. On the one hand they suffered considerably from frequent repetition (see Lecture XIV), and on the other hand any interruption in the work caused them to weaken or to disappear for a time altogether. So soon as complete differentiation between such compound stimuli had been established, the individual components when tested singly were found to have lost all their initial positive conditioned effect.
From a purely physiological point of view, the study of differentiation between a compound stimulus and its modification, which both contain the same elements but in s different order of succession, is of considerable interest. The experiments show that a compound stimulus the component units of which remain in themselves unaltered, and consequently most probably affect the same cells of the cerebral cortex, behaves in different modifications as a different stimulus, evoking in these cells now an excitatory process and now an inhibitory one. Plainly the experiments reveal the great importance of the synthesizing activity of the cortical cells which are undergoing excitation. These cells must form, under the conditions of a given experiment, a very complicated excitatory [p. 148] unit, which is functionally identical with the simple excitatory units existing in the case of more elementary conditioned reflexes. Such active cortical cells must necessarily influence one another and interact with one another, as has clearly been demonstrated in the case of compound simultaneous stimuli. The mutual interaction between the excited or inhibited cortical elements in the case of compound successive stimuli is more complicated; the effect of an active cortical cell upon the one next excited varies according to the influence to which it was itself subjected by the cell last stimulated. In this way it is seen that the order in which a given group of stimuli taking part in a stimulatory compound are arranged, and the pauses between them are the factors which determine the final result of the stimulation, and therefore most probably the form of the reaction, and we know already that different intensities of the same stimulus can be differentiated very accurately, one definite intensity being connected with excitation and another with inhibition.
It is evident from the description given in the present lecture that we must distinguish in animals an elementary, from a higher, type of analysis and synthesis. The former, and especially the elementary type of analysis, is based principally upon the properties and activity of the peripheral receptors of the analysers, while the latter is based principally upon those of the central ends of the analysers. Conditioned reflexes afford the means of investigating experimentally the functions both of the peripheral and of the central parts of the analysers, and such investigations have been conducted in our laboratories. The examples to be given during the remainder of this lecture will illustrate the scope of this field of animal experimentation.
In the first example I shall describe, it was sought to obtain by means of conditioned reflexes experimental data regarding the resonance theory of Helmholtz. We conceived that by partial destruction of the organ of Corti the disappearance of conditioned reflexes to certain tones would possibly be obtained. The following experiment was conducted by Dr. Andréev, who is still working on this subject. Pure tones were employed, being produced by two sets of apparatus, one giving tones from 100 to 3,000 and the other from 3,000 to 26,000 double vibrations per second. Various conditioned alimentary reflexes were established in the dog, namely, to tactile stimuli, visual stimuli, and different auditory stimuli (sound of a [p. 149] buzzer, metronome, a noise, and numerous pure tones). The cochlea was first completely destroyed on one side. When tested for the first time, six days after the operation, all the auditory conditioned reflexes were found to be present. A second operation (10th March, 1923) was now performed on the cochlea of the other side with the object of excluding only the lower part of the tonic scale. The osseous part of the cochlea was opened at the junction of its middle and upper thirds, and the exposed part of the membranous cochlea with the organ of Corti was injured with a fine needle. Already on the tenth day after the operation all the auditory stimuli, excepting tones of 600 double vibrations per second and lower, were found to be fully effective. In the course of three months following the operation, however, the effect of tones from 600 to 300 double vibrations became gradually restored. From numerous tests carried out from this period up to two years after the operation, the upper limit of the tones that had disappeared was fixed its somewhere between 309 and 317 double vibrations per second. The lower limit could not be determined, since we had no pure tones below 100 double vibrations per second at our disposal.
The following two tables are taken from the final period of the investigation:
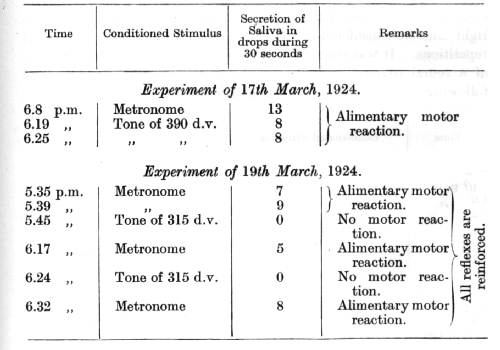 [p. 150]
[p. 150]
A histological examination of the cochlea has not yet been carried out, the dog still being used for experiments. It is evident, however, that the positive results of our experiments successfully replace the negative results obtained by Kalischer[1] with regard to this question.
Another problem which engaged our attention was whether the participation of both hemispheres was necessary for differentiation of sounds by their direction. This question was solved by the experiments of Dr. Bikov. The corpus callosum was severed in a dog, and after the animal had recovered from the operation the establishment of new conditioned alimentary reflexes was begun. Their formation presented no special difficulties, and proceeded at the same rate as in normal animals. One of the reflexes was established to the sound of a whistle, 1,500 double vibrations per second. The whistle, which was placed in a cardboard case, was supported on the wall on a level with the left ear and at a definite distance from the dog. The reflex appeared at the eighth repetition, and attained a maximum and permanent strength after 70 repetitions. The whistle was then transferred to the right side of the dog, and in this position was not reinforced by the unconditioned reflex. By repeatedly contrasting the sound from the left with that from the right, a differentiation of the direction of the sound was attempted. There was, however, not the slightest sign of any differentiation, even after 115 applications of the non-reinforced stimulus from the right, and we considered it futile to continue beyond this number of repetitions. It was concluded that a differentiation of the direction of a sound required a united activity of both hemispheres. The following is one of the most recent experiments :
 [p. 151]
[p. 151]
Other differentiations were obtained in this dog easily and rapidly, but never one involving localization of the source of a sound. In normal dogs the differentiation of sounds by their direction presents no more difficulty than any other differentiation, and is capable of great precision.
The experiments described in this and the preceding lectures leave no doubt in my mind that all the questions which have hitherto been considered as belonging to the domain of the so-called physiology of the organs of special sense can actually be investigated objectively by the method of conditioned reflexes. Are not Helmholtz's famous "unconscious conclusions"-- in his Physiological Optics -- in reality conditioned reflexes? We may take as an example the case of a drawing imitating the visual character of a relief. In actual experience, of course, the tactile and muscular stimuli proceeding from a relief represent the initial and fundamental stimuli : the visual stimuli provided by its areas of light and shade form the signalling conditioned stimuli, which only subsequently obtain a vital significance by being constantly reinforced by the tactile and muscular stimuli. In the further course of our lectures we shall refer to other examples which can be studied objectively in dogs, and which correspond fully with phenomena usually described only in connection with the physiology of the organs of special sense.
Footnote
[1] O. Kalischer. "Weitere Mitteilung über die Ergebnisse der Dressur als physiol. Unterschungsmethode auf den Gebieten des Gehör-, Geruchs- und Farbensinns." Archiv f. Anatomie und Physiologie, Physiologische Abteilung, p. 303, 1909.